

动物营养档案
卷64,第2号,2010年4月,121-135
单宁对猪生长性能和肠道生态系统的影响。
断奶仔猪
贾科莫·比亚吉(Giacomo Biagi)一个*,艾琳(Irene Cipollini)一个,布里吉特·R·保利克斯b 和弗朗兹·罗斯b
一个兽医和动物科学部门(DIMORFIPA),大学
意大利Ozzano Emilia的di Bologna;b动物营养与生产生理学系
慕尼黑工业大学,德国弗赖辛-魏恩史蒂芬
(2009年5月23日收到; 2009年10月2日接受)
单宁酸是天然的多酚化合物,可降低
饮食中的蛋白质还要表现出抗菌作用。本研究
在体外和体内研究了不同水平单宁的影响(使用
板栗木提取物
75%单宁)对生长性能的影响,
仔猪肠道菌群和壁的形态。在体外24小时内
盲肠发酵,单宁的利用率分别为0.75、1.5、3和6 g / l
减少了天然气的总产量以及氨和挥发性脂肪的浓度
酸和增加的肠球菌和大肠菌数量。当喂给
仔猪单宁分别为1.13、2.25和4.5 g / kg,可显着提高饲料效率
并降低了盲肠中氨,异丁酸和异戊酸的浓度。
空肠中的丹宁酸可能会增加乳酸菌的活力,
而细菌盲肠计数不受影响。回肠隐窝的深度倾向于
饲喂单宁的仔猪的含量降低了2.25和4.5 g / kg。本研究表明
用富含单宁的木材提取物喂养断奶仔猪可以改善
饲料效率和减少肠道细菌蛋白水解反应。的
单宁对低氧条件下肠球菌和大肠菌的生长促进作用
体外条件值得进一步研究。
关键词:盲肠微生物群性能;猪单宁酸
1.简介
单宁是在许多蔬菜中发现的天然多酚化合物
饲料,可以从几棵树的木材中提取
(库马尔和
Vaithiyanathan 1990)。单宁因其有害作用而闻名
将富含单宁的饲料喂食非动物饲料时,会对动物的生长性能产生影响
反刍动物。实际上,单宁酸会降低膳食蛋白的消化率(
Landın等。 2004年),因为它们能够与两种饮食形成不溶性复合物
蛋白质(巴特勒等,1984)和消化酶(詹斯曼等,1994)。除了他们
单宁还具有有益的抗营养作用(Ahn et al。
1998年; Min等。 2007)和止泻药(Palombo 2006)。在最初的几周
断奶后,由于仔猪经常腹泻和体重增加低
胃肠道和免疫系统的相对不成熟(Hedemann和
詹森
2004)。因为细菌引起的腹泻是
*通讯作者。电子邮件:giacomo.biagi@unibo.it
ISSN 1745-039X打印/ ISSN 1477-2817在线
2010泰勒和弗朗西斯
DOI:10.1080 / 17450390903461584

122
G.Biagi等。
仔猪肠道中有害细菌的繁殖,这个问题是
与可能作为副作用选择的抗生素和植物生长素抵消
肠道菌群中的抗生素抗性基因可能转移至
人类病原体(Phillips et al。2004)。由于这些担忧,使用
自欧盟以来,一直禁止使用抗生素促进生长
2006年1月,因此增加了对非药理学替代品的需求
调节年幼动物的肠道菌群。膳食补充
单宁可能减少仔猪腹泻的发生率,但提取物的作用
单宁酸对动物生长性能提出了一些关注。的目标
目前的研究是调查不同水平的单宁对生长的影响
猪盲肠菌群的体外反应,代谢及饲喂效果
单宁对仔猪生长性能,肠道菌群和壁形态的影响
体内。
2。材料和方法
2.1。动物护理和使用
在兽医的监督下进行动物饲养和护理
巴伐利亚政府办公室。处理方案确保适当的照顾和
按照德国动物保护法对所有动物进行处理。
2.2。体外研究
体外研究是在动物吗啡醇部门进行的
意大利博洛尼亚大学,动物学与动物生产(DIMORFIPA),之后
Biagi等人所述的方法。 (2006)。
预先消化猪的日粮(57%玉米,16%大麦和14%豆粕)
如Vervaeke等人所述在体外模拟回肠消化。 (1989)。这是一个
两步操作,其中饲料首先在胃蛋白酶-HCl溶液中孵育,然后在
胰酶溶液。预先消化的饮食在体外用作底物
发酵研究(Biagi等,2006)。六头猪(10个月大,
收集了160公斤活重,饲喂商业玉米-大麦-大豆饮食)
宰杀后,用缓冲液稀释(比例1:2; McDougall 1948)并过滤。的
过滤后的液体用作接种物。将接种物分配到五个10毫升中
玻璃注射器(每个注射器5毫升接种物)和五个50毫升容器(25毫升
每次处理,每个容器中接种一次,含20和100 mg预先消化的
饮食
(用作对照饮食)。密封注射器和血管,
在398℃温育24小时。
有五种治疗方法:对照饮食和对照饮食加单宁
(法玛坦1,斯洛文尼亚Tanin Sevnica),0.75、1.5、3.0和6.0 g / l。所有单宁
处理时,在孵育开始时,在密封注射器和
船只。将接种物的pH调节至6.7。
Farmatan是板栗(Castanea sativa mill)木材提取物(通过水获得)
提取),其中含有75%单宁,主要是没食子单宁(欧洲食品安全
管理局[EFSA] 2005)。半乳单宁是形成的可水解单宁
来自与糖的OH基相连的没食子酸(和鞣花酸)(特别是
葡萄糖)。 Farmatan的其他成分包括水(6.3%),鞣花酸(13.2%),
没食子酸(1.7%),单糖(2.6%),粗蛋白(0.8%),矿物质
(1.0%)和粗纤维(0.2%)。

动物营养档案
123
如Menke等所述测量产气量。 (1979),使用10毫升
玻璃注射器并记录每30分钟产生的累积气体量。
在发酵的6和24小时从每个容器收集发酵液样品。
孵育pH和氨气,并于24小时孵育挥发性脂肪酸(VFA)和
细菌计数的可行计数。
2.3。体内喂养研究
体内喂养试验是在动物营养和预防医学部进行的。
德国慕尼黑工业大学,生产生理学。四十八
杂交仔猪
(德国Landrace 6 Pietrain)在
28天和
从猪场运到谷仓里,他们被单独安置
网箱在受控环境中试用28天。适应4天后
所有仔猪均接受相同基础饮食的时期,动物(8.23 + 0.93公斤
BW)分为四组(每组12只动物),
体重,性别和垃圾。猪接受以下基本饮食:(i)不添加(对照)
饮食),或添加单宁(法马坦1)(ii)1.13 g / kg,(iii)2.25 g / kg,
和(iv)
4.5克/千克。所有饮食的配方均提供相同量的
能量,蛋白质,必需氨基酸,钙和磷。无抗菌
代理商被添加到饮食中。随意提供饲料和水
基础。实验饮食的成分和化学分析报告于
表格1。
分别称重动物并记录每只动物的饲料消耗
猪每周一次。每天记录浪费的饲料量。动物健康原为
在整个试验过程中进行监控。
在第28天,每次电击杀死6只动物,随后进行电击
完全出血。死亡后20分钟内,肠内容物(全
空肠和盲肠的内容以及回肠的最后100厘米的含量
表1.体内喂养试验中所用基础饮食的组成。*
成分
含量[g / kg]
化学成分
内容
棒子面
305
干物质{ [克/公斤]
885
小麦粉
273
粗蛋白质{ [克/公斤]
190
大麦粉
171
脂肪{ [克/公斤]
33.9
大豆粉47% CP
161
粗纤维{ [克/公斤]
32.0
豆油
10
钙[g / kg]
8.5
马铃薯蛋白
40
磷[g / kg]
6.0
碳酸钙3
5.7
赖氨酸[g / kg]
12.0
钙(H2PO4)2
5.7
蛋氨酸[g / kg]
3.9
预混料#
20
蛋氨酸 þ 半胱氨酸[g / kg]
7.2
盐酸赖氨酸
6.6
苏氨酸[g / kg]
7.8
DL-蛋氨酸
1.0
色氨酸[g / kg]
2.4
苏氨酸
1.0
ME [MJ / kg]
13.5
色p
0.2
注:*按日;获得了实验性日粮,将每公斤日粮中的0、1.5、3和6克玉米粉替换为
等量的法玛坦1 (含75%单宁)。#每千克饮食提供:钙6.16克; P,1.68g; m / z。
钠1.40克;镁0.28克;维生素A,16,800 IU;维生素D3,1,680 IU;维生素E,56毫克;维生素K3,1毫克;
硫胺素1毫克;核黄素3.5毫克;吡rid醇,2.1毫克;维生素B12,0.03毫克;烟酸16.8毫克;
d-泛酸
8.4毫克;叶酸,
2.8毫克;生物素
0.22毫克;胆碱,
420毫克;铁
140毫克;
锌140 mg;铜,28毫克;锰56 mg;我1.68毫克;硒,0.36毫克。{通过分析确定。

124
G.Biagi等。
分别收集)和粘膜(样本距离标本150厘米远)
幽门,在盲肠盲肠瓣之前100 cm处,在盲肠的顶端
对空肠,回肠和盲肠分别进行pH和氨采样)
测定和用于肠粘膜形态分析,而VFA
仅在盲肠样品中测定。空肠和盲肠的样本是
还培养了可行的细菌数量。
2.4。饲料,粪便样品,发酵液和肠道的化学分析
内容
饮食(CP,粗纤维和乙醚提取物)的分析根据
根据官方分析化学家协会的标准方法(AOAC 2000;
CP的方法954.01,粗纤维的方法962.09和乙醚的方法920.39
提取)。
发酵液和肠食糜中的氨含量根据
塞西等。 (1967)使用商业工具包(Urea / BUN – Color,BioSystems SA,
西班牙巴塞罗那)。用于测定肠食糜,食糜中的VFA
用蒸馏水按1:2的比例稀释,并离心(14,000 g,10分钟)和1 ml
将上清液装入微量离心管中,并用50 ml高氯酸脱蛋白
酸(默克,德国达姆施塔特)。 3小时后,将样品再次离心
(14,000 g,10分钟)。用于测定发酵液中的VFA,样品
离心
(3,000克,
15分钟)。上清液中VFA的浓度
发酵液和消化液通过气相色谱法测定(Biagi等。
2006).
2.5。细菌计数
收集食糜样品后,立即用9 ml稀释1 g样品
制备1%蛋白ept溶液并匀浆。食糜中的活菌数
(n ¼ 6)和发酵液(n ¼ 5)通过电镀10-
将稀释倍数的稀释液(在1%蛋白ept溶液中)加到乳酸杆菌培养基III琼脂板上
(638,德国DSMZ)用于乳酸菌,Difco DRCM琼脂平板
(Beckton,Dickinson and Company,Franklin Lakes,NJ,美国)梭状芽胞杆菌,Azide
用于肠球菌的麦芽糖琼脂平板(Biolife,Milano,意大利)和MacConkey琼脂
盘子
(N.
1.05465,默克,达姆施塔特,德国)。乳杆菌
将III型培养基和DRCM琼脂平板在398°C下孵育48小时
厌氧条件
(H2 þ 大约4至10% CO2; BBL GasPak加号
厌氧系统信封,贝克顿,迪金森公司,火花,医学博士,
美国)。将叠氮化物麦芽糖琼脂和MacConkey琼脂平板在室温下孵育24小时
在有氧条件下398C。
2.6。形态评估
空肠,回肠和肠黏膜标本的绒毛高度和隐窝深度
如Biagi等所述评估盲肠。 (2006)。固定粘膜样本
在10%中缓冲卡森的福尔马林并包埋在石蜡中;组织学切片
从组织块获得3 mm,垂直于粘膜表面切开,
用苏木精和曙红染色。组织形态计量学
使用计算机辅助图像分析系统执行
(Cytometrica,比克

动物营养档案
125
Gulden,意大利米兰)评估10个绒毛的高度(盲肠除外,其中的绒毛是
在随机选择的显微镜下,每节10个隐窝的深度和深度)
领域。
2.7。统计分析
2.7.1。体外发酵
使用改良的Gompertz细菌生长模型(Zwietering等,1992)进行拟合
天然气产量数据。该模型假设底物水平限制了
对数关系(Schofield等,1994)如下:
V ¼ VF 经验值F 经验值½1 ðð米米Ë=VFÞðltÞg;
其中V ¼ 在时间t,t产生的气体量 ¼ 发酵时间,VF ¼
产生的最大气体量,m米 ¼ 最大产气率,其中
发生在气体曲线的拐点处 ¼ 滞后时间,作为时间-
在拐点处切线的轴线截距(Zwietering等,1990)。
使用程序GraphPad Prism 4.0(GraphPad
软件,美国加利福尼亚州圣地亚哥)。使用GLM通过ANOVA分析数据
完全随机化的SAS(SAS Inst。,Inc.,Cary,NC,USA)程序
设计。线性和二次对比用于确定图像的性质。
对单宁添加的反应。每个注射器和器皿形成
试点。差异被认为具有统计学显着性(p 5 0.05)。
2.7.2。体内喂养试验
使用SAS(SAS Inst。,Inc.
完全随机的设计。线性和二次对比
用于确定对单宁进食反应的性质。
每个仔猪组成实验单元。统计上考虑差异
在p 5 0.05时具有显着性。
3.结果
3.1。体外实验
产气曲线
(数字
1)被修改准确描述
Gompertz模型(r2 ¼ 0.92)。天然气产量结果列于表2。
单宁酸降低了产量(线性,p 5 0.05),而最大气体产生率
单宁容器中的产量趋于增加(p ¼ 0.06).
孵育6小时后,单宁酸(线性,
p 5 0.001;表2),但补充丹宁酸对24小时的pH值没有影响。
6 h后单宁可显着降低氨气浓度(线性
和二次方,p 5 0.001;表3),且减少幅度为730%
将单宁以0.75 g / l添加到740%,单宁以6 g / l使用。 24小时后
在培养过程中,单宁可以显着减少氨(线性和二次
p 5 0.001),单宁浓度分别为0.75 g / l和6 g / l,从732%到763%。
温育24小时后,与对照相比,乙酸减少了
单宁酸
(线性和二次方,p 5 0.01;表格
3)和最低的乙酸
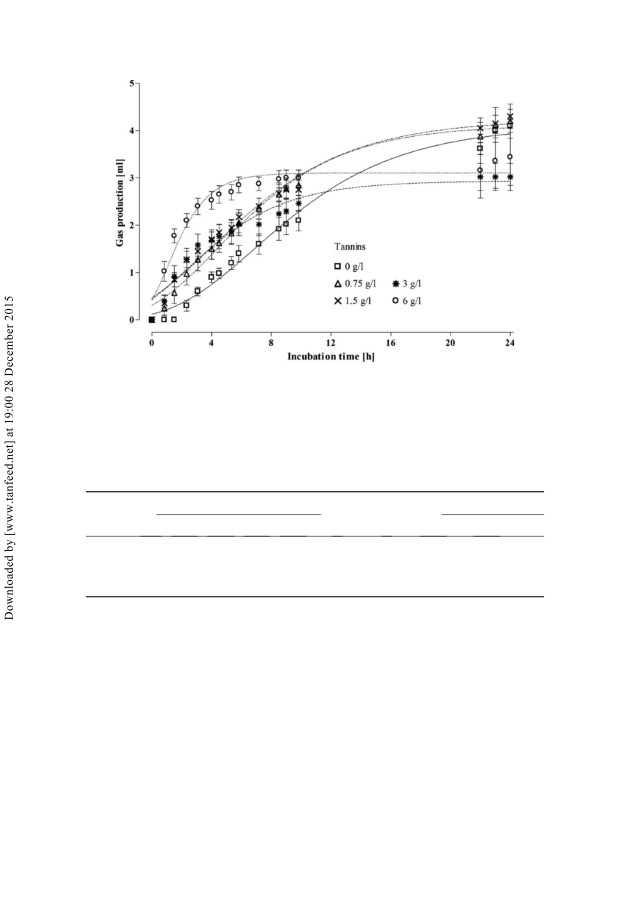
126
G.Biagi等。
图1.猪盲肠接种物与或一起体外孵育后的产气曲线
无需添加不同浓度的单宁。用
每种处理均单独使用五个注射器,以适应改良的Gompertz生长模型。手段
绘制曲线,条表示SEM。
表2.修正的Gompertz方程式适合于24小时内的产气量数据和pH
猪盲肠接种物与不同浓度单宁的体外温育
添加单宁[g / kg]
对比度,p
汇集
的p
0
0.75
1.5
3
6
扫描电镜模型
线性二次
VF*
4.37
4.25
4.24
2.91
3.86
0.264
0.011
0.031
0.487
米m#
0.29
0.30
0.31
0.31
0.48
0.037
0.056
0.013
0.069
6小时的pH
7.03
6.99
6.94
6.96
6.87
0.017
50.001
50.001
0.865
24小时的pH值
6.80
6.80
6.82
6.86
6.81
0.030
0.711
0.472
0.679
注意:* VF,产生的最大气体量[ml];#米米,最大产气速率[ml / h]。价值观
是每种饮食中五次重复的最小二乘均值。
浓度确定为每升6克单宁酸(716%)。丙酸(线性
和二次方,p 5 0.05)和正丁酸(线性和二次方,p 5 0.01)
单宁从0.75降低到4-12%和从8-36%显着降低
和6克/升。异丁酸和异戊酸的浓度为
单宁显着降低(线性和二次方,p 5 0.001),并且这种降低
单宁分别为0.75和6 g / l的范围从21-52%和29-57%。
单宁显着降低了挥发性脂肪酸的总浓度
(线性,p 5 0.001)。
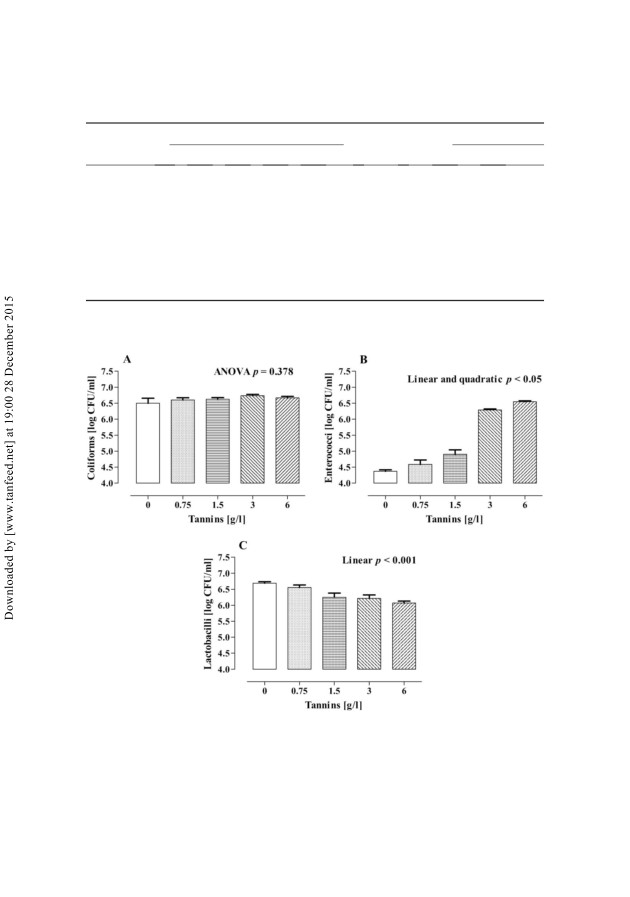
图2和图3表示6和24小时时细菌的存活数,
分别。在6小时时,大肠菌的存活数量不受治疗影响;
相反,孵育24小时后,大肠菌群增加(线性和二次方,
动物营养档案
127
表3.氨(6和24小时后)和挥发性脂肪酸的浓度[mmol / l]
不同浓度的猪盲肠接种物的体外培养(24小时后)
单宁
添加单宁[g / kg]
对比度,p
汇集
的p
0
0.75
1.5
3
6
扫描电镜模型
线性的
二次方的
氨
在6小时
12.4
8.66
8.12
7.97
7.37
0.232
50.001
50.001
50.001
在24小时
20.6
14.1
9.50
8.91
7.72
0.435
50.001
50.001
50.001
醋酸
42.8
44.2
41.8
38.0
36.1
0.511
50.001
50.001
50.01
丙酸
16.9
16.3
15.4
13.9
15.0
0.267
50.001
50.001
50.05
异丁酸
1.22
0.96
0.84
0.67
0.58
0.009
50.001
50.001
50.001
正丁酸
9.10
8.39
7.71
7.28
5.81
0.088
50.001
50.001
50.01
异戊酸
1.03
0.73
0.63
0.50
0.45
0.013
50.001
50.001
50.001
C2 / C3 *
2.53
2.71
2.72
2.74
2.41
0.004
50.001
0.159
50.001
正C4 / isoC4#
7.47
8.72
9.17
10.8
9.95
0.010
50.001
50.001
50.001
总酸
72.4
71.7
67.5
61.4
58.9
0.792
50.001
50.001
0.117
注释:* C2 / C3,乙酸与丙酸之比;#n-C4 / isoC4,正丁酸与异丁酸之比。值是
每种饮食的最小二乘均值是五次重复。
图2.大肠菌群6小时后的大肠菌群(A),肠球菌(B)和乳杆菌(C)的活菌计数
不同浓度单宁的猪盲肠接种物的体外培养
表示每次处理+ 5个血管的SEM)。
p 5 0.01)所添加容器中丹宁酸的大肠菌浓度最高
单宁3 g / l(þ0.5 log CFU / ml)。乳酸菌明显减少
单宁添加量和乳酸菌最低计数确定为6克单宁
128
G.Biagi等。
图3.在24小时的大肠菌群(A),肠球菌(B)和乳杆菌(C)的活菌计数
猪盲肠接种物与不同浓度单宁的体外温育
表示每次处理+ 5个血管的SEM)。
每升(孵育6小时和24小时后分别为70.7和70.8 log CFU / ml);
线性,P 5 0.01)。单宁显着增加了肠球菌计数,
单宁最高含量为6克/升(þ2.2和 þ6和24小时后2.5 log CFU / ml
孵化;线性和二次方,p 5 0.05)。梭状芽孢杆菌的可行计数
不受治疗影响,在6和24小时后平均为7.7和7.8 log CFU / ml
分别进行孵育(数据未显示)。
3.2。体内喂养试验
动物活重,平均日增重和每日饲料摄入量均不显着
单宁影响
(表4)。整个饲料效率
试验28天
单宁酸(线性,p 5 0.05)显着改善,饲料效率最低
在饲喂单宁为4.5 g / kg的猪中观察到这种现象(75%;表4)。
实验饮食对肠道pH没有显着影响。平均pH值
空肠,回肠和盲肠分别为5.70、6.53和5.77(数据
显示)。盲肠氨浓度呈下降趋势。
饲喂单宁的猪(p ¼ 0.07;表5),但未观察到氨水平的影响
从空肠和回肠的食糜样品中。
盲肠食糜中丙酸的浓度呈上升趋势。
单宁喂养猪的减少(p ¼ 0.06;表5)。减少仔猪的单宁喂养

动物营养档案
129
表4.添加或不添加饮食的仔猪的生长性能
断奶后四个星期内的单宁。
添加单宁[g / kg]
对比度,p
汇集
的p
0
1.13
2.25
4.5
扫描电镜模型
线性二次方
最终体重[kg]
19.3
19.4
19.7
20.7
0.680
0.478
0.152
0.549
日增重[g / d]
395
400
410
445
17.30
0.183
0.045
0.388
ADFI [克/天]
549
553
570
588
23.79
0.638
0.214
0.752
进料效率
1.39
1.38
1.39
1.32
0.019
0.034
0.023
0.105
注意:数值是每种测试日粮中12头仔猪的最小二乘均值。
表5.空肠,回肠和盲肠的食糜中氨的浓度[mmol / l]
补充饮食的仔猪盲肠的食糜中的食糜中的挥发性脂肪酸和
在断奶后的四个星期内没有不同浓度的单宁。
添加单宁[g / kg]
对比度,p
汇集
的p
0
1.13
2.25
4.5
扫描电镜模型
线性二次方
氨
空肠
12.6
14.2
12.8
12.3
1.201
0.721
0.725
0.420
回肠
20.2
16.7
17.7
16.5
1.788
0.497
0.270
0.553
盲肠
37.6
27.5
30.6
25.4
3.022
0.071
0.039
0.449
醋酸
74.1
61.5
75.2
65.6
4.695
0.151
0.590
0.305
丙酸
43.4
35.2
45.8
36.0
3.009
0.058
0.428
0.809
异丁酸
1.56
0.88
1.01
0.71
0.141
0.003
0.002
0.212
正丁酸*
17.5
13.7
20.8
14.0
1.385
0.006
0.613
0.314
异戊酸
1.53
0.87
0.93
0.72
0.124
0.001
50.001
0.095
总酸*
142
116
148
119
7.526
0.016
0.298
0.892
注意:值是每种测试日粮六头仔猪的最小二乘均值。 *立方(p 5 0.01)
比较0至4.5 g / kg的单宁酸。
(线性,p 5 0.01)盲肠的异丁酸浓度(从735%到755%
单宁分别为2.25和4.5 g / kg)和异戊酸(从739%至
单宁含量分别为2.25和4.5 g / kg的753%)。饮食疗法也有
对正丁酸和总VFA浓度有显着影响(立方,p 5 0.01)。
空肠和盲肠食糜中细菌的存活计数表示为
图4.单宁喂养倾向(p ¼ 0.06),以增加乳酸杆菌的存活数
在空肠中,盲肠乳酸菌不受饮食治疗的影响。
空肠中大肠菌群,肠球菌和梭状芽孢杆菌的浓度低于
检测极限,无法确定。大肠菌的盲肠计数显示
猪饲喂丹宁酸时增加的趋势(p ¼ 0.08),而盲肠
梭菌和肠球菌不受单宁补充的影响。单宁
补充对空肠和回肠的绒毛长度没有影响(表6),但是
显着影响回肠隐窝深度(立方,p 5 0.05)。
4。讨论
在本试验中,单宁的利用并未显着影响动物
当以4.5 g / kg的日粮饲喂单宁时,体重增加但饲料效率提高。
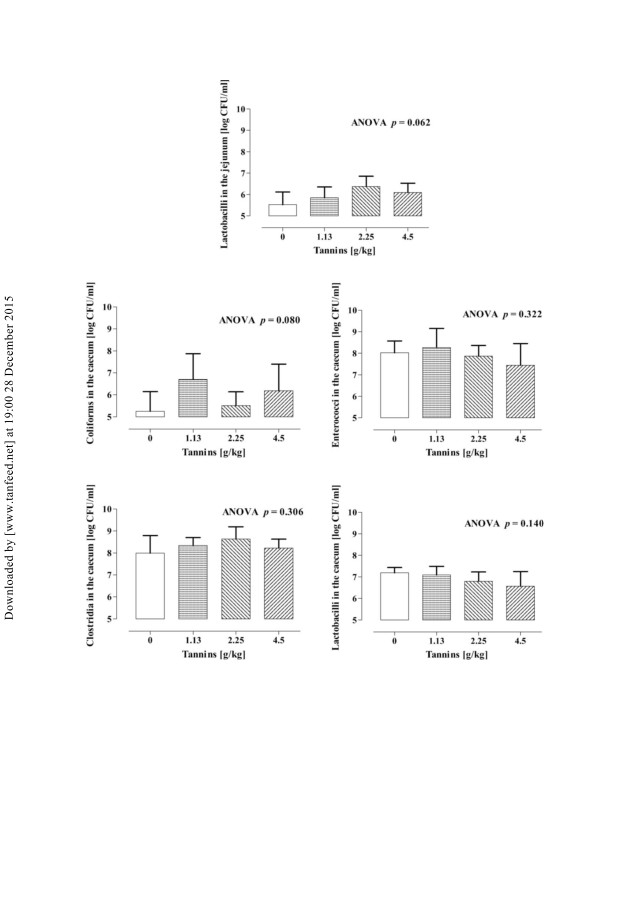
130
G.Biagi等。
图4.空肠中大肠菌,肠球菌,梭菌和乳杆菌的活菌计数
饲喂不同浓度单宁的仔猪的盲肠和食糜(值是平均值+ SEM
每次处理中有六只动物)。
饲喂高水平单宁的仔猪日粮的利用率提高
有争议的发现,因为单宁酸已被认为是抗营养因子
它们降低膳食蛋白质消化率的能力(Mariscal-Landın等人,2004)。在
Lizardo等人的研究。 (1995年),在猪日粮中添加了单宁含量丰富的
高粱导致动物生长性能下降。相反,其他作者
观察到以高丹宁酸为原料的猪饲料,例如田豆(Flis

动物营养档案
131
表6.仔猪肠道黏膜的绒毛高度和隐窝深度[mm]
断奶后的四个星期中,饮食中添加或不添加单宁水平不同的食物。
添加单宁[g / kg]
对比度,p
汇集
的p
0
1.13
2.25
4.5
扫描电镜
模型
线性的
二次方的
空肠
绒毛
413
466
442
466
26.78
0.457
0.272
0.589
地穴
356
396
358
360
31.01
0.768
0.859
0.557
回肠
绒毛
313
328
297
314
27.11
0.881
0.806
0.969
地穴*
272
310
226
241
19.74
0.033
0.059
0.587
盲肠
地穴
348
425
348
358
25.22
0.120
0.666
0.197
注意:值是每种测试日粮六头仔猪的最小二乘均值。 *立方(p 5 0.05)
比较0至4.5 g / kg的单宁酸。
等。 1999年)和角豆粉(Lizardo等,2002年)不影响动物的生长。在一个
断奶猪的研究(Myrie等人,2008年),以15 g / kg的单宁喂养动物
尽管回肠
苏氨酸的表观消化率降低了,但总蛋白的消化率却没有降低。朗斯塔夫
和McNab(1991)观察到,低剂量的富含丹宁酸的蚕豆能增强大豆的抗逆性。
雏鸡空肠和回肠消化物中脂肪酶的活性什么时候
据报道,饲喂高粱的单胃动物的生长性能下降
(Lizardo等,1995),蚕豆(Rubio等,1990),羽扇豆(Kim等,2007)
与其他单宁含量丰富的饲料相比,
传统的猪饲料,例如大豆粉和玉米粉,这些饲料
通常以淀粉的消化率较低为特征
(鲁尼和
Pflugfelder 1986; Wiseman 2006)和蛋白质(Mariscal-Landınet al。2002)部分,
以及存在大量其他抗营养因子(Huisman
等。 (1990)。
在目前的体外研究中,单宁酸对维生素的代谢有很大的影响。
猪盲肠微生物群。实际上,单宁浓度呈线性增加
减少了总的气体产量并降低了乙酸,丙酸和正丁酸的浓度
酸,对盲肠细菌的活性有抑制作用。因为
乙酸与丙酸的比例不受单宁添加的线性影响,似乎
单宁会降低总细菌活性,而不有利于特定代谢
途径。相反,单宁浓度的增加导致线性
增加正丁酸与异丁酸的比例。这一发现可能由以下原因解释
产生丁酸的细菌的碳水化合物代谢相对较高,
以及异丁酸的还原,异丁酸是蛋白质的代谢产物
细菌分解代谢。
有趣的是,单宁含量为4.5 g / kg决定了最高的产气量,
这表明,尽管具有一般的抑制作用,但某些细菌的活性还是
高水平的单宁选择性刺激。实际上,肠球菌的计数(以及
单宁显着增加大肠菌群的含量,而
乳酸菌计数减少。当饲喂仔猪时,单宁酸没有增加
肠球菌盲肠计数,但趋于增加空肠中的乳酸菌和
盲肠中的大肠菌。已经显示出几种肠道细菌菌株

132
G.Biagi等。
可以降解单宁酸,没食子酸和鞣花酸,并将其用作能源(Bhat
等。 1998年;塞尔达(Cerda)等人。 2005年)。尽管单糖含量相对较低
(2.6%)的木材提取物,也可能含有一些提取物
在体外发挥作用但无法达到的肠球菌生长因子
单宁饲喂仔猪时的动物后肠,因此无法增加
肠球菌盲肠计数。在体外,肠球菌生长增强可能解释
由于对底物的竞争加剧,乳杆菌数略有减少。
然而,更高的肠球菌计数可能仅仅是更高的结果
与乳酸菌相比,肠球菌的生长潜力。
单宁的定义是指一组水溶性多酚
分类为可水解和不可水解(缩合)单宁(Akiyama et al。
2001)。单宁化合物的抗菌活性不同(Akiyama等人,2001;
Puupponen-Pimia等。 2005年),单宁的组成受到植物的影响
的来源和提取方法。单宁对体外的抗菌作用
大肠埃希菌的致病性菌株(Yao等,2006),产气荚膜梭菌和
金黄色葡萄球菌(Ahn等,1998)和幽门螺杆菌(Funatogawa等,1998)。
(2004年),但在我们的研究中,任何浓度的单宁都没有
不能减少大肠菌群和梭状芽胞杆菌的体外计数。在Ahn等人的研究中。
(1998年),单宁从荆芥中获得,并显示出对
MIC值为0.5-8 mg / ml的不同细菌菌株
与我们在本试验中使用的单宁浓度相当。
缺乏针对大肠埃希菌的大肠菌和梭状芽胞杆菌的体外抗菌活性。
体内研究证实了我们使用的单宁。目前没有数据
关于在小肠吸收的膳食单宁的量
猪,因此很难估计有多少单宁会到达动物身上
后肠。对单宁喂养大鼠的研究表明,单宁酸主要是
在粪便中回收,但在中发现少量单宁化合物
血浆和尿液样本,表明单宁吸收发生了(Clifford
和布朗(2005)。目前,尚无证据表明丹宁酸可能会参与其中。
动物肠道具有与已观察到的相同的有效抗菌作用
一些作者(Ahn等,1998; Funatogawa等,2004; Yao等,2006)
体外条件。
氨,异丁酸和异戊酸的浓度为
显着降低体内和体外,表明单宁有效
减少细菌蛋白水解反应。实际上,上述异酸是由
缬氨酸和亮氨酸的去氨反应(Van Soest 1982),具有指示性
氨氮对细菌蛋白的代谢程度。氨是一种有毒的化合物,
可以破坏细胞并降低绒毛高度(Nousiainen 1991),并且一旦被吸收
必须在损失能量的情况下将其转化为尿素(Eisemann和Nienaber,1990年)。
此外,亚急性浓度的氨可能会降低动物的生产能力
(Visek 1978)。几种膳食补充剂已显示可减少肠道
细菌蛋白水解:其中包括不可消化的低聚糖和益生菌
(Piva et al。2005),有机酸(Roth and Kirchgessner 1998)和草药提取物
(Ushida等人,2002)。根据目前的结果,单宁似乎是有价值的
替代上述饮食补充剂以控制肠道
断奶猪的细菌蛋白水解反应。
补充单宁对肠粘膜组织学的影响也
调查。早期断奶对肠粘膜有巨大的负面影响
仔猪的形态(Gu et al。2002),可能导致养分吸收不良

动物营养档案
133
(Boudry et al。2004)。正丁酸是粘膜的首选能量底物
回肠(Chapman et al。1995)和大肠(Roediger 1980),
观察到用丁酸钠喂养仔猪(Galfi and Bokori 1990; Wang
等。 2005)对肠道粘膜有积极的营养作用,增加了
回肠微绒毛的长度和盲肠隐窝的深度。相反,其他作者
丁酸钠对肠粘膜无营养作用
(Biagi等人2007)或丁酸前体(Piva等人2002)喂给断奶仔猪
猪。在本研究中,单宁对盲肠正丁酸有立方效应
单宁浓度高时回肠隐窝的浓度和深度趋于减少
用量为2.25和4.5 g / kg。目前,我们还没有解释这种影响
单宁。不过,因为小肠中的隐窝主要有分泌物
功能(Wood 2006),减少其表面可能有助于降低严重程度
仔猪断奶后腹泻。
5.结论
目前的研究表明,断奶仔猪饲以单宁
丰富的木材提取物可以提高饲料效率并减少肠道
细菌蛋白水解反应。然而,增长促进作用
似乎对大肠菌群有木材提取物值得进一步研究。
参考文献
安YJ,李CO,权JH,安JW,朴JH。 1998. Galla的生长抑制作用
源自罗伊斯的单宁对肠细菌的作用。 J应用微生物学。 84:439-443。
秋山H,藤井K,山崎O,大野T,岩城K.2001。几种抗菌作用
单宁对金黄色葡萄球菌。抗微生物化学杂志。 48:487-491。
官方分析化学家协会(AOAC)。 2000。官方分析方法。 17日
ed。盖瑟斯堡(MD):霍维兹(W Horwitz)。
Bhat TK,Singh B,Sharma OP。 1998.单宁的微生物降解
– 一种趋势
透视。生物降解。 9:343-357。
Biagi G,Piva A,Moschini M,Vezzali E,Roth FX。 2006.葡萄糖酸对仔猪的影响
生长性能,肠道菌群和肠壁形态。 J动漫科学。
84:370-378.
Biagi G,Piva A,Moschini M,Vezzali E,Roth FX。 2007。性能,肠道菌群,
丁酸钠的断奶仔猪的体形和壁形态。 J动漫科学。 85:1184-1191。
Boudry G,Peron V,Huerou-Luron I,Lalles JP,Seve B.2004。断奶会同时导致短暂
并长期改善仔猪的吸收,分泌和屏障特性
肠。 J食品134:2256-2262。
巴特勒L,里德尔DJ,莱布里克DG,布莱特HJ。 1984年。蛋白质与高粱的相互作用
单宁:机制,特异性和重要性。 J Assoc Off Chem Soc。 61:916-920。
塞尔达(Cerda B),佩里亚戈(Periago P),埃斯金(EspınJC),托马斯-巴伯兰(Tomas-Barberan) 2005.尿石素a鉴定为
人结肠菌群由鞣花酸和相关化合物产生的代谢产物。
J农业食品化学。 53:5571-5576。
查普曼,格拉恩MF,哈顿M,威廉姆斯NS。 1995年。
溃疡性结肠炎患者的末端回肠粘膜。英国外科医师82:36-38。
克利福德(MN),布朗(美国)。
2005。类黄酮:化学,生物化学及其应用。
第6章:膳食类黄酮与健康-拓宽视野。博卡拉顿(FL):
CRC出版社/泰勒和弗朗西斯。 p。 331-370。
Eisemann JH,JA Nienaber。 1990.进食和禁食的组织和全身氧气吸收
引导。英国J食品。 64:399-411。
EFSA(欧洲食品安全局)。 2005.添加剂科学小组的意见
以及动物饲料中使用的产品或物质,应委员会的要求
Farmatan产品对兔子和小猪的安全性和有效性。 EFSA J.222:1-20。

134
G.Biagi等。
Flis M,Sobotka W,Purwin C,Zdunczyk Z.1999。含野外饮食的营养价值
猪的原花青素含量高或低的大豆(Vicia faba L.)种子。 J动画供稿
科学8:171-180。
船川K,林S,下村H,吉田T,波多野T,伊藤H,平井Y.2004。
药用植物来源的可水解丹宁酸的抗菌活性
幽门螺杆菌。微生物免疫。 48:251-261。
Galfi P,Bokori J.,1990年。《用正丁酸钠饮食的猪的饲养试验》。 Acta
兽医38:3-17
顾欣,李丹,佘荣。2002。断奶对仔猪小肠结构和功能的影响
小猪。拱动画食品。 56:275-286。
Hedemann女士,Jensen BB。 2004.胃和胰腺中酶活性的变化
断奶仔猪周围的组织和消化道。拱动画食品。 58:47-59。
豪斯曼(Huisman J),范德波尔(van der Poel)空军基地,韦斯特根MWA,范韦登(van Weerden EJ)。 1990年。抗营养
猪生产中的因素(ANF)。 World Rev Anim产品。 25:77-82。
Jansman AJM,Enting H,Verstegen MWA,Huisman J. 1994年。缩合单宁的影响
蚕豆壳(Vicia faba L.)对食糜中胰蛋白酶和胰凝乳蛋白酶活性的影响
从猪的小肠收集的。英国J食品。 71:627-641。
Kim JC,Pluske JR,Mullan BP。 2007。羽扇豆作为猪饲料中的蛋白质来源。 CAB版本
透视农业兽医科学营养资源。 2(编号003):12。
Kumar R,Vaithiyanathan S.,1990年。动物的发生,营养意义及其对动物的影响
树叶中单宁的生产力。动画Feed科学技术。 30:21-38。
Lizardo R,Canellas J,Mas F,Torrallardona D,Brufau J. 2002年。角豆粉的利用
仔猪日粮中的维生素及其对断奶后生长性能和健康的影响。 34emes
法国皇家骑师协会(Journees de la Recherche Porcine),法国动物科学协会会员。
法国巴黎。 p。 5-7。 Fevrier 2002。
Lizardo R,Peiniau J,Aumaitre A.1995。高粱对蛋鸡生产性能,消化率的影响
断奶仔猪的膳食组成和胰,肠酶活性
小猪。动画Feed科学技术。 56:67-82。
Longstaff MA,McNab JM。 1991年。浓缩单宁含量丰富的豆壳的影响(蚕豆
faba L.)对食糜中脂肪酶(EC 3.1.1.3)和α-淀粉酶(EC 3.2.1.1)的活性
和胰腺以及幼雏对脂质和淀粉的消化作用。英国J食品。
66:139-147.
Mariscal-LandınG,Avellaneda JH,Reis de Souza TC,Aguilera A,Borbolla GA,3月B。
2004.高粱中的单宁对氨基酸回肠消化率和胰蛋白酶的影响
生长猪的胰凝乳蛋白酶活性。动画Feed科学技术。 117:245-264。
Mariscal-LandınG,Lebreton Y,Seve B.
2002年。表观和标准化的真正回肠
全部提供的蚕豆,羽扇豆和豌豆的蛋白质和氨基酸的消化率
猪饲料中去皮或挤出的种子。动画Feed科学技术。 97:183-198。
McDougall EI。 1948年。反刍动物唾液研究。 1.绵羊的组成和产量
唾液。生物化学杂志43:99-109。
Menke KH,Raab L,Salewski A,Steingass H,Fritz H,Schneider W. 1979年。
反刍动物饲料中气体的消化率和代谢能含量
将它们与瘤胃液一起在体外孵育时的产量。农业科学杂志。 93:217-222。
Min ER,Pinchak WE,Anderson RC,Callaway TR。 2007.单宁对体外的影响
大肠杆菌O157:H7的生长和普通大肠杆菌的体内生长
从ste牛皮。 J食品保护。 70:543-550。
Myrie SB,Bertolo RF,Sauer WC,Ball RO。 2008.常见抗营养因子和
猪日粮中的纤维性饲料对氨基酸消化率的影响,特别是
苏氨酸。 J动漫科学。 86:609-619。
Nousiainen J. 1991.益生菌和奥拉喹多作为饲料的比较观察
断奶仔猪的添加剂。 2.对绒毛长度和隐窝深度的影响
空肠,回肠,盲肠和结肠。 J动画生理动物食品。 66:224-230。
Palombo EA。 2006.用于治疗的传统药用植物的植物化学物质
腹泻:作用方式和对肠道功能的影响。 Phytother水库。
20:717-724.
菲利普斯一世,凯斯韦尔M,考克斯T,德格鲁特B,弗里斯C,琼斯R,夜莺C,普雷斯顿R,
Waddell J.2004。在食用动物中使用抗生素是否会对人类健康构成威胁?
对已发布数据的严格审查。抗微生物化学杂志。 53:28-52。

动物营养档案
135
Piva A,Luchansky JB,Biagi G.2005。乳糖醇,乳酸菌或其混合物的影响
结合(合生元)对体外肠蛋白水解和断奶仔猪饲料效率的影响
猪。可以J Anim Sci。 85:345-353。
Piva A,Prandini A,Fiorentini L,Morlacchini M,Galvano F,Luchansky JB。 2002年。
和乳糖醇协同增强肠粘膜的营养状态;
降低保育猪肠道中的组胺水平。 J动漫科学。 80:670-680。
Puupponen-Pimia R,Nohynek L,Alakomi HL,Oksman-Caldentey KM。 2005年。
浆果酚类抗人肠道病原体。生物因子。 23:243-251。
Roediger WE。 1980.厌氧细菌在结肠粘膜代谢福利中的作用
在人中。肠道。 21:793-798。
Rooney LW,Pflugfelder RL。 1986年。特别强调影响淀粉消化率的因素
在高粱和玉米上。 J动漫科学。 63:1607-1623。
Roth FX,Kirchgessner M.1998。有机酸作为幼猪的饲料添加剂:营养
和胃肠道作用。 J动画Feed科学。 7(增补1):25-33。
Rubio LA,Brenes A,Castano M.,1990年。生的和蒸煮的蚕豆的利用
(Vicia faba L.,var。minor)和用于种鸡的日粮中的蚕豆成分。
英国J食品。 63:419-430。
Schofield P,Pitt RE和Pell AN。 1994.体外气体生产中纤维消化的动力学。
J动漫科学。 72:2980-2991。
Searcy RL,Reardon JE,工头JA。 1967年。一种新的光度法测定血清尿素
氮测定。我是J Med技术。 33:15-20。
Ushida K,Maekawa M,Arakawa T.2002。饮食中草药的影响
提取物对猪大肠中挥发性硫的产生。食品科学维生素J。 48:18-23。
Van Soest PJ。 1982年。反刍动物的营养生态学。科瓦利斯(OR):O和B书。
p。 162。
Vervaeke IJ,Dierick NA,Demeyer DL,Decuypere JA。 1989年。精力充沛
猪中纤维消化的重要性。二。后肠消化的实验方法。
动画Feed科学技术。 23:169-194。
Visek WJ。 1978年。通过氨调节饮食和细胞生长。我是J临床食品。 31(Suppl.10):
216-220.
王建峰,陈玉霞,王志新,董世SH,赖志伟。 2005。丁酸钠的影响
断奶仔猪小肠黏膜上皮的结构中国兽医科学
技术。 35:298-301。
Wiseman J.2006。非反刍动物中淀粉消化率的变化。动画Feed科学技术。
130:66-77.
伍德JD。 2006。胃肠道生理学。章节
41:病理生理
肠易激综合征的基础学。阿姆斯特丹:爱思唯尔学术出版社。
p。
1009-1032.
姚K,何Q,应佳D,石冰。2006。精制紫荆丹宁提取物的潜力。纳特
产品资源。 20:271-278。
Zwietering MH,Jongenburger L,Rombouts FM,van't Riet K.,1990年。细菌建模
增长曲线。应用环境微生物。 56:1875-1881。
Zwietering MH,Rombouts FM,van't Riet K.1992。滞后定义的比较
细菌生长的阶段和指数阶段。 J Appl细菌。 72:139-145。