
其中PB是结合蛋白的量(g / kg DM),而PPP是植物可沉淀的酚类化合物(%DM)。在线性反应和平稳线相交处发生的饮食PPP临界百分比为6.4%。PPP混合物结合的最大蛋白质量估计为37.24 g蛋白质/ kg DM。缩合的单宁酸占蛋白质结合蛋白观察值变化的81%。
众所周知,CT夹杂物可降低瘤胃CP降解的速度和程度(Waghorn,2008; Patra和Saxena,2011)。 莲花角ic 是一种经常被研究的含CT的草料,经常被评估为苜蓿的替代品 。莲花 角线虫(0.972.77%CT)线性降低了立即可溶的CP分数和原位降解率 ,因此与紫花苜蓿 (0%CT)相比,降低了牧草RDP的比例 (Williams等,2010 ; Coblentz和Grabber,2013)。蛋白质降解程度在饲草CT浓度上的下降趋势表明,每单位CT可保护平均0.61单位的CP免受瘤胃降解的影响(Coblentz和Grabber,2013年)。
其他有效的含CT饲料包括 Lespedeza stuevei Nutt。和 A. angustissima var。 hirta 被鉴定为 以1:1的比例混合时对瘤胃豆粕蛋白体外降解具有极高保护作用的植物 (Johnson等,2015)。在同一研究中,CP在48 h降解的程度与六个暖季多年生豆类中可沉淀蛋白质的多酚浓度成反比(Johnson等,2015)。
当补充CT提取物时,观察到相似的结果。 葡萄 (葡萄)种子提取物(GSE)与羽扇豆(Lupinus angustifolius var )复合 。 Tanjil 种子CP(分别为96和180 mg GSE / g CP的比例)降低了美利奴羊公羊的速溶级分和CP降解率,从而导致RUP的比例随着CT浓度的增加而增加(Bruno-Soares等,2011)。 。然而,CP降解的有效降低与CT的两倍增加不成比例(Bruno-Soares等,2011)。Quebracho单宁提取物(QTE)是来自Schinopsis的 CT来源 spp。,持续报道降低多种膳食纤维(Dschaak等,2011)和CP水平(Aguerre等,2016)的瘤胃CP降解。响应似乎与剂量相关,因为从1%到6%的DM内含物增加,线性降低瘤胃CP降解(Ahnert等,2015)。
由于添加CT引起的瘤胃蛋白质降解减少,随后降低了NH 3 浓度,为动物提供了进一步的价值。超过微生物生长需求的瘤胃中可用氮以NH 3的形式被吸收,在肝脏中代谢为尿素,并在尿中循环或排泄。高蛋白降解率会增加尿中N的排泄量,这会对环境产生不利影响(Powell等,2010),降低N的利用效率,并产生与尿素合成和排泄增加相关的代谢负担,从而阻碍动物的生长(Van Duinkerken等) (2005年; Kohn等人,2005年)和生育率(Westwood等人,2000年; Tshuma等,2014)。
通过CT夹杂物增加RUP会持续降低 多种物种在体外 和 体内的瘤胃NH 3 -N浓度 ,除非饲料蛋白质水平远高于动物的需求。通常观察到剂量反应,并且与CT的蛋白质沉淀能力有关(Johnson等,2015)。在连续培养模型中,用 日粮L. corniculatus 干草(总日粮CT的0.32和0.97%)代替乳制品配比中的一半或全部 紫花苜蓿干草 可使 NH 3 -N浓度和流量分别降低至少15%和25% (Williams et al。,2010)。CT浓度较高 在 奶牛口粮中, 当L. corniculatus 青贮饲料(日粮总CT的7.3-9.5%)代替苜蓿青贮饲料(日粮总CT的0%) 时,体内 瘤胃NH 3浓度降低了24% (Hymes-Fecht等, 2013)。然而,一些研究报道CT对瘤胃NH 3 -N浓度没有影响(Williams等,2011; Dickhoefer等,2016)。这些结果可以用CT源,较低的水摄入量(即 像Dickhoefer等人所述的较低的NH 3稀释度)来 解释。(2016年) ]或因为能源可用性限制了微生物的生长。 Christensen等。(2015年) 还报道了 用L. corniculatus (0.51%CT)代替50%或100%的苜蓿干草 后,泌乳奶牛瘤胃NH 3 -N浓度 无差异。缺乏反应似乎是由于变异较高(1.244 mg / 100 mL)所致,因为苜蓿草 日粮(8.33 mg / 100 mL)与混合饲料(6.05 mg / 100 mL)中的NH 3 -N浓度较高 以及 基于角质乳酸杆菌的 饮食(6.70 mg / 100 mL)(Christensen等人,2015)。但是,在鸟足三叶形饮食中微生物蛋白质的生产得以改善,这表明当提供含CT的草料替代苜蓿时,可以更有效地利用饮食中的氮。作为瘤胃NH 3 尿液中的CT移位N减少(Deaville等,2010 ; Williams等,2011 ; Hymes-Fecht等,2013 ; Ahnert等,2015 ; Orlandi等,2015 ; Aguerre等人,2016),可通过减少粪便中的一氧化二氮排放量来显着提高反刍动物生产的环境可持续性(Powell等人,2010)。
与对瘤胃CP降解和NH 3 浓度的公认反应相比,CT对微生物蛋白质合成和微生物生长效率的影响尚不清楚。在DM的1,2,和3%的QTE除了被报道提高羊饲喂微生物效率 紫花苜蓿 干草(铝Dobaib,2009年)和 格塔丘等。(2008年) 报道仅在浓度为0.5%和1%时有所改善,而在1.5%处没有。相比之下,在DM摄入量的2、4、6%处注入QTE 会使十二指肠微生物蛋白流量减少11%,21%和39%(Dickhoefer等人,2016年),而当QTE的浓度大于10%时,其添加量可高达36%。 DM摄入量的1%(Ahnert et al。,2015)。
对于不同的动物物种,补充含CT的饲料似乎具有相似的矛盾反应。对于奶牛而言,饲喂角质乳 而不是 苜蓿可以 增加微生物蛋白的生产(Christensen等,2015)。相反, 使用多种膳食CT浓度(0-200 g / kg DM)进行的多个 体内绵羊试验(n = 11)报告对微生物蛋白质合成没有影响(Min等人,2003年)。由于微生物蛋白对反刍动物氨基酸需求的重要贡献,因此对含CT的饲草或提取物的未来评估应着眼于对微生物蛋白生产和效率的影响。
Waghorn等人的工作基于利用CT改善反刍动物N代谢的最初兴趣 。(1987),他报告说,由于L. corniculatus CT,瘤胃后必需氨基酸的通量增加了50%, 随后肠内氨基酸的平均利用率提高了60%。这些结果的大小尚未复制。通常,由于较大的RUP比例,CT增加了瘤胃后的氨基酸通量,但对肠氨基酸利用率的影响差异很大。除了饮食组成以及动物的生理状态和生产水平外,反应似乎还取决于CT来源和化学特性(Kariuki和Norton,2008年)。
根据结合亲和力,大多数CT-蛋白质复合物被认为在厌恶的酸性条件下会解离,从而将两种化合物释放到消化基质中(Patra和Saxena,2011; Hagerman,2012)。小肠中性pH条件为CT-养分结合提供了另一个机会,尽管随着pH升高至中性以上,复合物形成的可能性较小(Hagerman等,1992)。CT-蛋白质相互作用的亲和力和结合强度会影响整个消化道的蛋白质消化率。
Kariuki和Norton(2008) 直接证实了瘤胃后绵羊中CT蛋白质的解离。牛血清白蛋白作为与CT的复合物通过一个烟气导管引入时,具有超过82%的真实消化率,这表明大部分CT结合蛋白已被释放并在瘤胃后可用。然而, 角质乳杆菌 CT改善了绵羊的瘤胃后氨基酸供应,但是降低了肠内氨基酸的利用率( Waghorn等,1994b)。但是,血液中的氨基酸浓度表明,由于CT导致流向肠道的N流量增加,从而弥补了氨基酸消化率降低的趋势;因此,净吸收没有受到影响( Waghorn等,1994b)。
向霍尔斯坦ste牛 添加9-17 g / kg DM的相思木鞣质提取物可使进入十二指肠的氨基酸通量平均比对照治疗的平均值高30%(Orlandi等人,2015)。对于乳制品生产而言,最重要的是蛋氨酸和糖异生氨基酸供应受CT补充的积极影响。随着CT夹杂物含量的增加,表观和真实的N消化率均线性降低7%或以下,而N保留量则从17.3线性提高至33.2 g / d(Orlandi等,2015)。这一发现表明,瘤胃后氨基酸供应的改善超过了蛋白质消化率的任何损害。这些数据也为CT对内源性蛋白质分泌的影响提供了一些见识。
内源性蛋白质损失的增加(主要是通过增加粘液的产生(Sell等人,1984年)或与消化酶的CT复合作用(Al-Mamary等人,2001年))经常被认为是两种反刍动物肠道蛋白质消化率受损的原因。 (Waghorn,2008)和非反刍动物(Jansman,1993)。 Orlandi等。(2015年) 报道了按明显的基础计算,以及仅通过使用中性洗涤剂不溶性氮作为膳食来源氮的替代物来消除内源性损失的影响时,氮消化率的抑制程度相似。这表明 相思树 DM浓度为0.9-1.7%的单宁酸不会刺激内源性蛋白质分泌。CT是否增加内源蛋白的分泌,明显降低了总管蛋白的消化率,总体氮保留量和动物生产能力,是评估CT补充效果最重要的指标。
在假设的CT-蛋白质相互作用的背景下,CT的命运仍不清楚。当潜在的稳定的CT蛋白复合物从瘤胃转移到厌恶菌及其他地方时,会发生什么变化?一种假设是,在厌恶中遇到酸性pH时,黄酮键之间的酸水解会解聚CT。但是,如果瘤胃或异胃中的CT未被降解,则CT可能会与任何未消化的蛋白质,肽或氨基酸重新复合,从而阻止这些化合物在小肠中吸收并减少氨基酸的潜在增加被动物吸收的酸。这可能是从尿中N排泄转变的一个可能的解释(Grainger et al。,2009反刍动物食用富含CT的饮食时,尿素酶活性降低,粪便中氨气挥发(Aguerre等,2011)。
体内 实验可以帮助回答与动物生产中CT功效有关的更多实际问题。每天饲喂DM量为1-6%的QTE添加了草料和浓缩饲料的完全成长的小母牛降低了表观总道CP消化率,剂量大于2%,但是当QTE大于4%时,其他营养物质的消化率受到损害。 (Ahnert et al。,2015)。尽管消化率具有负面影响,但与对照组相比,QTE夹杂物的总氮保留量均得到改善,而与剂量水平无关。该结果表明,只要有足够水平的可消化能量,肌肉的生长就不会受到损害。Aguerre等在泌乳的荷斯坦奶牛中观察到了对总养分消化率的类似响应 。(2016年),据报道线性消化率下降是由于在DM的0.45%,0.90和1.80%处加入了单宁混合物(1/3栗提取物,2/3 quebracho提取物)。牛奶生产效率(kg牛奶/ DMI)得到改善,但牛奶蛋白产量(kg真蛋白/天)呈线性下降。
动物农业必须有效利用饲料中的氮,并防止氮过多地释放到环境中,以实现并保持可持续性。浓缩单宁酸是营养学家实现这一目标的潜在工具。尽管需要澄清CT对瘤胃后氨基酸供应和内源N分泌的影响,但公认的是,低至中等的膳食CT浓度会减慢瘤胃中CP的降解,从而导致RUP比例增加和瘤胃后氨基酸通量增加。保守地假设,由于表观蛋白质消化率降低,肠氨基酸丰度的改善被否定,反刍动物氮平衡和生理的改善似乎主要归因于瘤胃NH 3的减少 浓度,因此减少了与尿素排泄和低效氮循环相关的代谢成本。此外,CT代表了一个机会,可以控制饮食中的RUP并针对瘤胃后氨基酸的特性,以更紧密地匹配动物的需求并提高牛肉和奶制品的生产效率。
单宁缩合的驱虫活性
反刍动物生产者最担心的问题之一是胃肠道线虫寄生虫,尤其是 捻转血矛线虫。 与这些感染相关的主要成本包括使用驱虫药治疗,因动物产量下降而造成的损失以及因动物死亡而造成的损失,所有这些都因寄生虫种群的驱虫抗药性增加而加剧(Leathwick和Besier,2014年)。向反刍动物提供含有CT的饲料或饲料可能会减轻胃肠道寄生虫问题。在波尔山羊中, 在50%和75%的DM浓度下添加 Lespedeza cuneata可使粪卵数,特别是 捻转血Haemonchus contortus减少84.6%和91.9%(Terrill等,2009)。)。 Minho等。(2008) 报道 了当 给予CT molesima molissima(15%CT,DM基础)的CT提取物时,绵羊的 臭气中粪便卵数和成年的Haemonchus contortus的减少 ,但未观察到对 小肠成年的 Trichostrongylus colubriformis的影响。CT的驱虫特性不仅可能是寄生虫特有的,而且还可能是隔室特有的(Tedeschi等人,2014年):一些人报道了CT可以治疗厌恶症而不是小肠中的胃肠道线虫寄生虫。
使用含CT的植物作为驱虫药的主要挑战是,并非所有含CT的草料都具有驱虫特性(Naumann等人,2014a)。来自Leucaena retusa,Lespedeza stuevei 和 Acacia angustissima var的浓缩单宁 。 hirta 减少了扭纹梭菌的幼虫迁移 ,分别降低 了65.4%,63.1%和42.2%。来自Desmanthus illinoensis,Lespedeza cuneata 和 A. angustissima的浓缩单宁酸 对L3幼虫的效力较低。像CT-蛋白质相互作用一样,响应于不同CT来源的驱虫活性变化可能是结构-功能关系。 Quijada等。(2015年) 结果表明,主要由原花青素亚基组成的CT的驱虫活性要高于主要由原花青素组成的CT。这可以解释为什么 L.藤, 主要的prodelphinidin亚基组成,其产生CT草料(瑙曼等人,2015B。 ,已经证明高驱虫效力) 体内 (Shaik等人,2006。 ; 特里尔等人,2007。 ; 特里尔等人,2009)。牛寄生虫也有类似的发现。缩合的单宁主要由原蛋白原素组成,降低了L1的活力,而成年期的 Ostertagia ostertagi 和 Cooperia oncophora (Desrues等人,2016年)。
有足够的证据表明,较小的CT聚合物比较大的CT聚合物具有更高的驱虫活性(Naumann等,2014a, Barrau等,2005)。但是, Quijada等。(2015) 和 Desrues等人。(2016年) 报道了相反的效果,并建议结果的变化可能是由于CT提取和纯化的差异和困难,以及不同CT对胃肠道线虫寄生虫的作用方式的差异。CT对胃肠道线虫寄生虫的作用方式尚未完全阐明。然而,用CT提取物孵育后,线虫幼虫的电子显微照片显示线虫表皮发生了改变,并且CT结合在了头部区域(敌人的人,2012)。
缩合单宁与其他植物专用代谢产物的结合可能会影响这些化合物作为驱虫药的功效。 Klongsiriwet等。(2015) 证明了结合类黄酮如槲皮素和木犀草素与CT的协同作用。黄酮类化合物的存在下,使用体外 幼虫体外试验可以增强CT的驱虫活性 。
使用HT作为潜在的驱虫药也有希望。 Engström等。(2016年) 表明,许多HT几乎没有驱虫效果。但是,那些主要由PGG结构组成的蛋白表现出对卵孵化和幼虫运动性的抑制作用。有证据表明,在存在HT的情况下,卵,幼虫中体和头侧区域的外部结构会发生变化(Engström等,2016)。
矿物和缩合单宁之间的相互作用
单宁和其他多酚除了具有与蛋白质结合并使其沉淀的能力外,还可以有效地与铁结合,并在较小程度上与铜,锰,铝,锌和钴结合。多酚在多大程度上金属螯合会影响矿物质的生物利用度。但是,反刍动物和单胃动物中补充CT的评估很少涉及对矿物质营养的潜在影响。尽管CT对体内矿物质利用率的影响 仍然不明确,但人们对该现象背后的化学原理有了更好的了解。矿物螯合的最低要求是在苯环上至少存在两个相邻的羟基(邻位或邻二羟基苯基)(Andjelkovic等,2006))。这种化学结构有助于植物的色素沉着以及整个植物环境中的阳离子养分循环(Quideau et al。,2011)。概括地说,CT分子中的邻二羟基苯基官能团越多,金属螯合的可能性就越大(McDonald等,1996)。相互作用是pH依赖性的,并且极大地影响CT的抗氧化能力(Kumamoto等,2001)。铁和铝阳离子在pH值为3.20或更低时与CT结合,而镁,钙,锌,铜,锰和钴阳离子在pH大于3.70时与CT结合(Faithfull,1984)。但是,通常认为这些复合物在很宽的pH范围内以及在整个胃肠道中都稳定(McDonald et al。,1996; M。 Kumamoto et al。,2001 ; Scalbert等,2002)。
由于具有强大的铁结合潜力,铁是与单宁有关的最常研究的矿物。早期研究酚类化合物对铁吸收的抑制作用的研究表明,膳食中单宁的添加量与抑制程度之间存在密切的联系(Brune等,1989)。从那时起,已经确定了对铁和其他矿物质结合很重要的官能团。具有邻苯二酚和没食子酰基的单宁是有效的金属螯合剂(Andjelkovic等,2006; Perron和Brumaghim,2009)。每个CT分子可结合两个或多个金属离子,每个金属离子可与来自两个不同CT分子的邻二羟基苯基形成螯合物 (McDonald等,1996)。Fe结合需要黄酮类B环上的3’,4′-二羟基基团(图6)(Khokhar和Owusu-Apenten,2003年),游离羟基的增加与Fe结合能力的提高有关(Andjelkovic等。 ,2006; Mladénka等人,2011)。
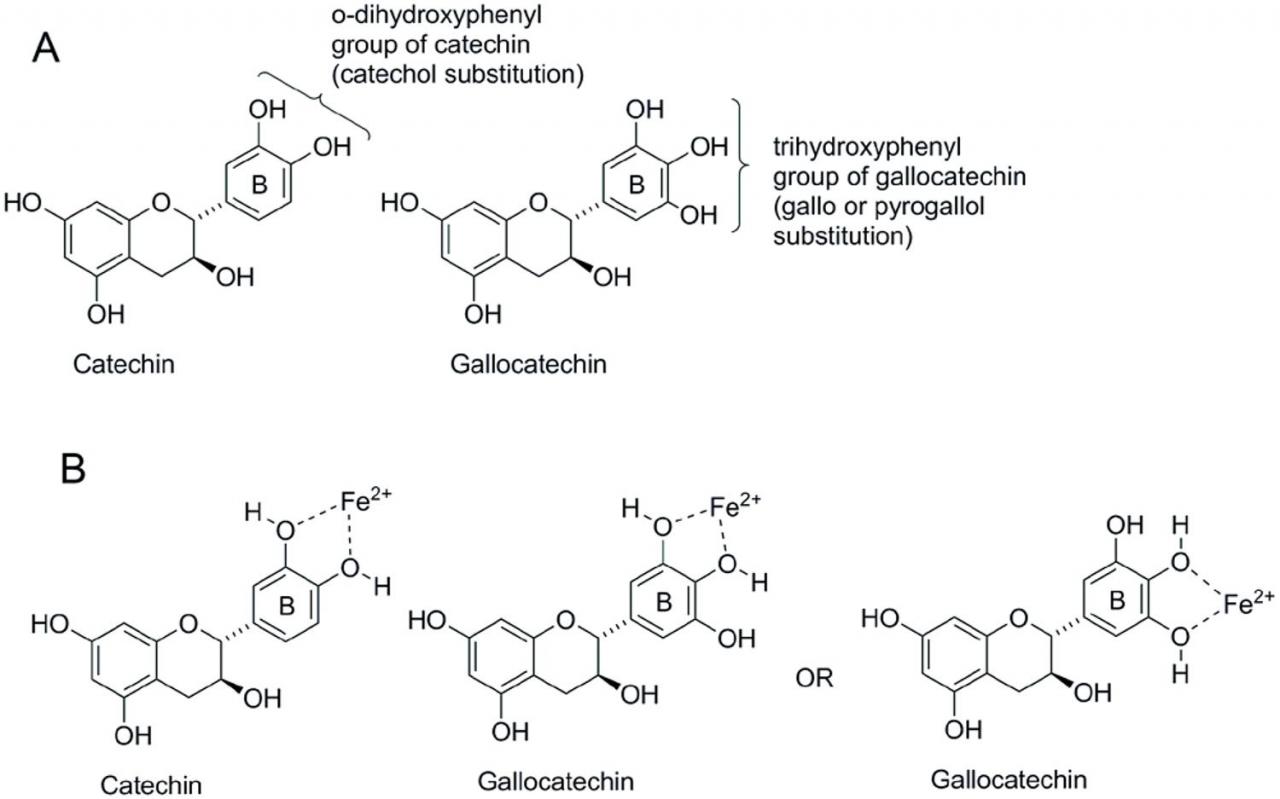
缩合的单宁铁复合物可以有效地抑制铁的吸收(Lavin等,2010; Wren等,2013),单宁酸的含量低至5 g / kg(Afsana等,2004)。在细胞培养中,单宁酸与铁的1:1比例抑制了92%的铁吸收(Glahn和Wortley,2002年)。当作为一顿饭的一部分用于更复杂的饲料基质中时,这种作用减弱了,但仍然很明显(Yun等人,2004年),表明矿物质吸收抑制的程度受消化过程中饲料基质中其他营养物质的存在影响。与CT形成的络合物相反,抗坏血酸和乙二胺四乙酸通过可逆或不可逆地与Fe形成更易溶的络合物来增强Fe的生物利用度。含有高抗坏血酸浓度成分的饮食即使在低浓度下也能增加铁的吸收并有效抑制CT-Fe的螯合(Tamilmani和Pandey,2016年)。
缩合的单宁酸通过相似的化学机理与铜结合。与其他阳离子一样,CT化学结构,尤其是羟基取代的类型,可以解释其螯合Cu的能力差异。邻苯三酚比邻苯二酚羟基化方式更有效地沉淀Cu,并且随着pH降低,沉淀减少(McDonald等,1996)。与先前的报道相反,茶单宁已显示可增加大鼠体内铜的生物利用度(Scalbert等,2002)。
锌对CT的亲和力比铁和铜低(McDonald等,1996),特别是在酸性和中性pH条件下(Santos-Buelga和Scalbert,2000)。 Afsana等。(2004) 证明补充单宁对活体内锌吸收没有影响 。 CT与其他阳离子复合物的报道很少,通常与茶的消费有关。据报道,茶单宁可以抑制人和大鼠的铝吸收(Fairweather-Tait等,1991),但在其他研究中却没有作用(Greger和Lyle,1988)。
除了直接结合外,阳离子还可能影响CT与膳食蛋白的相互作用。钙被发现可以增强茶单宁,表没食子儿茶素没食子酸酯和牛奶中的β-乳球蛋白的蛋白质结合能力,从而可以形成更大的CT-蛋白质复合物(Carnovale等人,2015)。CT对维生素吸收和代谢的影响尚不清楚。但是,这可能是一个重要的考虑因素,因为单宁酸已被证明会对大鼠的维生素A状况产生负面影响,并且可能与硫胺素相互作用并减少维生素B的吸收(Jansman,1993年)。
CT与金属离子的相互作用不仅影响矿物质的生物利用度,而且是CT在肠腔中作为有效抗氧化剂的机制之一(Brenes等,2008; Perron和Brumaghim,2009)。面对越来越多的动物生产中使用抗生素的限制,对CT改善肠道健康的兴趣迅速增加(Redondo等,2014)。但是,有关体内 CT,维生素和矿物质之间相互作用的信息 是缺乏。在一定程度上,只要不损害动物的性能,就可以假设矿物质状态是适当的。但是,由于用于CT的强铁螯合潜力以及可能发生的许多养分相互作用,用于动物生产的CT研究应考虑矿物质的平衡。
目前有关CT对反刍动物矿物质营养影响的报道非常不一致。通过研究 Waghorn等。(1994a),评估了CT 对羊的新鲜切花 莲梗(5.5%CT)的影响,报道了CT降低了S吸收并增加了P和Zn的净吸收,而对其他矿物质(Fe,Cu,Ca,P (和Mg)很小或不受影响。松树皮CT(DM的0.06-1.11%)线性降低了K,S和Cu的消化率,而肉山羊的P,Mg,Mn,Zn和Fe的消化率增加了(Min等,2015)。瘤胃中受CT束缚的某些矿物质有可能在小肠中获得。Pagan-Riestra等。(2010年)证明磷从瘤胃到小肠的消失与A. angustissima具有生物活性的CT有关 。
丹宁酸对单胃矿物质平衡的影响更为频繁,并且随着CT浓度的增加,矿物质的利用率通常会下降。食用日粮中GSE升高但水平较低(0.025、0.25、2.5和5 g / kg)的肉鸡的血浆铜,铁和锌的浓度线性下降(Chamorro等人,2013)。高鞣质高粱(1.36%CT)降低了肉鸡中Ca,P,Mg,Na,K,Fe和Co的表观吸收(Hassan等,2003)。同样,随着高粱单宁含量增加至日粮DM的3%,矿物质的消化率呈线性下降(Mahmood等,2014)。一般而言,鲜有关于在猪日粮中添加CT的报道,而对矿物质消化率影响的测量方法则更为罕见。单宁酸是一种可水解的单宁,已被证明可以降低断奶仔猪日粮中的铁利用率(Lee et al。,2010)。缩合的单宁酸已被有效地用于抑制多种外来物种中铁的吸收并减轻铁过载(Wood等,2003; Lavin等,2010; Lavin,2012)。
最后要考虑的是含CT成分对饮食的贡献。CT螯合金属的能力可以促进植物生长环境或日粮加工过程中金属的积累。在这种情况下,CT可能成为动物体内金属离子的贡献者,而不是降低金属离子作为金属离子螯合剂的利用率。这对于防止铁超负荷症或其他矿物质毒性是有问题的。随着CT和反刍动物相互作用的研究的进展,测量和报告CT对氮素代谢和总体表现以及矿物质平衡的影响将非常重要,以确保长期补充CT的安全性。
CT肠胃发酵相互作用
甲烷是一种有力的温室气体,通常在瘤胃的微生物发酵过程中产生,并在发酵过程中释放到环境中。牲畜产生的甲烷气体 是大气中CH 4的第二大来源,据估计占 美国人为CH 4排放总量的22%。牛肉和奶牛占这些排放量的96%(美国环保局,2016年)。然而,当以CO 2 当量表示时,畜牧业占温室气体总排放量的4.2%。牛肉和奶牛是 畜牧业生产CH 4的主要动力,仅占总贡献量的3.6%(USEPA,2016年)。
产甲烷是一个复杂的过程,瘤胃中的产甲烷酶将纤维素分解为动物可用的形式。 Buddle等。(2011年) 开发了此过程的简化示意图,其中瘤胃细菌,原生动物和真菌对进入瘤胃的饲料起作用。来自瘤胃微生物的代谢副产物包括H 2,CO 2和挥发性脂肪酸(VFA)等。 在代谢H 2的处理过程中会产生肠CH 4。在挥发性脂肪酸形成过程中没有消耗的还原当量可用于产生CH 4,这代表了动物代谢能的损失。
瘤胃中形成的许多重要的共生关联之一是产甲烷的古细菌与纤毛原生动物之间的关系(Ng等,2016)。已经提出这种结合是为了促进代谢H 2 从原生动物到产甲烷菌的种间转移。二氧化碳通过甲烷以产生能量减少,产生CH 4 作为以下面的方式代谢的副产物:4H 2 + CO 2 →CH 4 + 2H 2 O.它还假定ħ 2 结合CO 2 可以被同种乙酸原酶用来产生乙酸盐,乙酸盐是反刍动物的主要能源。因此,如果较少的H 2 转化为CH 4,则更多的H 2 可用于VFA生产,从而导致动物的代谢能增加。
向反刍动物饲喂含CT的饲料或饲料可能是减轻 反刍动物家畜CH 4排放并增加可代谢能量摄入的有效自然方法。多种具有不同类型CT的豆类物种在体外抑制CH 4 产生 (Naumann等,2013b)。但是,当将这些含CT的豆类作为唯一的饲料进行发酵时,它们也抑制了总产气和VFA的产生,表明消化率受到抑制。这表明抑制CH 4 不会导致H 2 从产甲烷菌和产甲烷作用转变为同型产乙酸菌和VFA的产生,并且证明了将CT用于CH的缺点4 缓解。目标应该是在 不影响产量的情况下减少反刍家畜的CH 4排放,这需要有选择地减少CH 4的 产生而又不减少总气体或发酵量。用CT可以达到这个目的。在 体外 瘤胃发酵研究中,用玉米苜蓿饲料的45%的饲草部分替代库氏 乳杆菌 或 沙棘D. ,分别使膳食CT浓度达到2.6%和9%,减少了CH 4的 产生而不降低总的天然气产量(Naumann等,2015b)。
CT影响产甲烷反应并减少 反刍动物产生CH 4的机制尚不十分清楚。关于CT如何抑制甲烷生成,有多种假设,但没有一个得到明确的证明。一种假设是CT直接作用于瘤胃中的产甲烷菌。 Ng等。(2016) 报道了一种基于蛋白质的粘附素的存在,该粘附素可能位于菌毛尖端,其在促进产甲烷菌-原生动物共生中起作用。细胞包膜的一部分,包括细胞膜,壁和糖萼,也含有蛋白质。CT可能会与这种蛋白质黏附素或部分细胞膜结合,从而干扰产甲烷菌-原生动物复合物的建立并减少种间H 2 传递。抑制这种共生关系也可能对纤毛原生动物种群产生负面影响。 Bhatta等。(2015) 决定的,当饲料从包含CT纤毛虫种群瘤胃降低 孟加拉榕 和 Azardirachta籼稻 在26和13.8%的浓度,分别。
另一个假设是通过降低营养物质对瘤胃微生物的利用率,从而降低底物的消化率并间接抑制瘤胃微生物种群而发生间接抑制。因为CT与矿物质(Lavin,2012)和有机分子(如蛋白质)(Saminathan等,2014),碳水化合物(Soares等,2012a)和脂质(Delehanty等,2007)结合,所以这些复合物不仅不能用作瘤胃微生物的底物,而且CT会结合到调节其活性的微生物酶上(Gonçalves等,2011)。 Naumann等。(2013c)表明CT结合的蛋白与CH 4减少之间的关系较弱。
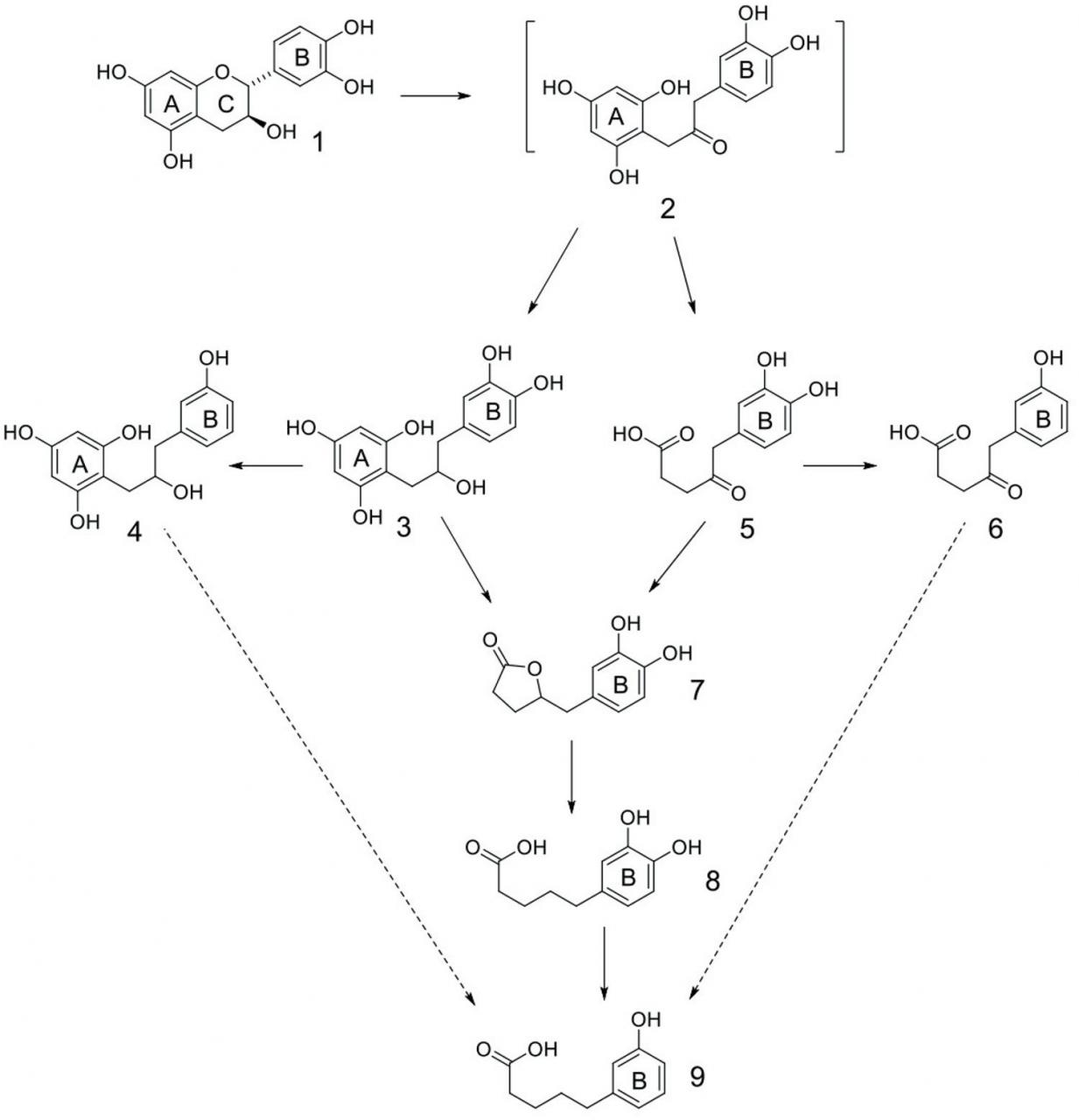
CT如何抑制CH 4的第三个假设 是CT充当氢沉(Naumann等人,2013a)。 贝克尔等。(2014年) 报道了在体外 瘤胃液环境中减少甲烷生成的 趋势,这种现象通过添加flavan-3-ol儿茶素线性发生。在该实验中,按照儿茶素分子多达六个氢原子被儿茶素的降解产物和CH捕获4 生产在1.2摩尔CH的速率降低4 每摩尔儿茶素。根据这项研究,作者报道了儿茶素作为氢原子受体(氢沉)而产生的代谢产物的特征。这些发现与人类微生物区系从flavan-3-ols的生物转化中分离出的化合物相似(Feng,2006; Monagas等,2010)。简短地说,儿茶素被转化为过渡中间体2,该中间体接受两个氢原子以提供还原的化合物3(图7)。随着C-4酚键的氢解发生,化合物3接受另外两个氢原子,将其还原成CH键并生成化合物4。化合物5可以通过断裂A环从瞬态中间体2衍生而来,切除乙酰乙酸分子。化合物5接受另外两个氢原子当量,这些氢原子当量经过B环的C-4酚基的脱羟基反应,得到化合物6。化合物3可以进行A环断裂,中间羟基酸从中进行内酯化以生成化合物7。化合物5中酮官能团的还原产生与3相同的中间体羟基酸,并提供内酯7。7中内酯的CO键的氢解需要两个氢原子当量,得到酚酸8。化合物8中的C-4酚键的氢解反应(再次需要两个氢原子当量)提供了酚酸9。尽管在本研究中未检测到中间体以支持其随后转化为化合物9,但化合物4可能会经历类似于那些的生物转化。提出将化合物3转化为化合物9(A-环断裂/内酯化,内酯氢解)以递送化合物9。最后,推测化合物6可以通过还原/内酯化,然后进行内酯氢解顺序而转化为化合物9。所有结构(化合物4可能会发生类似于化合物3转化为化合物9的生物转化(A环分裂/内酯化,内酯氢解)以递送化合物9的作用。最后,化合物6可能通过还原/内酯化,然后进行内酯氢解顺序。所有结构(化合物4可能会经历类似于化合物3转化为化合物9的生物转化(A环断裂/内酯化,内酯氢解)以递送化合物9。最后,化合物6可能通过还原/内酯化,然后进行内酯氢解顺序。所有结构(通过与市售材料进行比较来确认或通过质谱法和核磁共振波谱表征,除化合物2外的图7)。
预测瘤胃CH 4的 产生对于反刍动物的营养很重要,尤其是对于估计可代谢能量的摄入以评估动物可用于代谢的总总能量而言。无论是通过增加可代谢能量的摄入来提高动物生产效率的利益,还是为了努力对气候变化产生负面影响,减轻污染的需求都是有益的。预测 饮食成分对肠CH 4的抑制作用对于在不影响动物生产的前提下达到缓解或减少目标非常重要;畜牧业的目标是减少而不是完全抑制肠道CH 4。
已经开发出方程式,以 根据饮食成分或各种与动物相关的变量预测瘤胃CH 4的产生。例如, Dittmann等。(2014年) 开发了预测方程 ,使用线性回归描述了反刍动物和骆驼科动物体重与每日CH 4排放之间的关系。体重解释 了反刍动物在CH 4排放中观察到的变化的93%,骆驼科动物的91%。两种线性回归均显示出正相关,因此,随着体重增加,CH 4 排放增加。尽管相关性很强,但与这些方程式相关的挑战之一是,尚无法确定驱动CH的参与因素4个 变量。体重和CH 4之间关系的可能驱动因素之一 是干物质摄入(DMI)。随着动物体型的增加,DMI的数量也会增加,这可能会增加可发酵的有机物质,进而增加肠道CH 4的 产生。另一个挑战是将体重作为变量来缓解CH 4。这样做将导致选择较小的动物以减少CH 4的 排放。
Ramin和Huhtanen(2013) 开发了线性和二次方程式,用于 基于DMI预测每日肠内CH 4的产生。摄入量与CH 4 产生呈正相关。对于线性回归,DMI解释了CH 4 排放量变化的85%。但是,需要确定与肠CH 4 产生最密切相关的参与因子,才能将DMI应用于缓解工作。例如, Grainger和Beauchemin(2011) 开发了方程,用于 根据饮食脂肪预测牛和羊的肠CH 4产量。肠溶CH 4 产量与总膳食脂肪呈负相关。绵羊的预测方程具有比牛更大的负斜率,这表明膳食脂肪在减轻 绵羊体内的CH 4方面可能比牛更有效。
Mangino等。(2003) 开发了牛肠发酵模型来预测 基于总能量摄入的CH 4排放。有趣的是,该预测方程包括甲烷转化因子,该因子基于饲料总能量转化为CH 4的比例。转换因子特定于美国不同地区的饮食特征和动物类型。例如,加利福尼亚州的奶牛甲烷转化率为4.8,而西方的奶牛甲烷转化率为5.8。甲烷转化率仅在奶牛和小母牛身上有所不同,表明不同类别和种类的肉牛将总能量以相同的速率转化为甲烷,而与区域差异和动物类型无关。
尽管已经进行了大量研究来评估产生CT的草料降低肠内CH 4产生的潜力 ,但很少有研究集中于 根据饲料CT含量预测对CH 4排放的抑制。 Naumann等。(2013b)评估了CT,VFA和肠CH 4 产生之间的关系。由于增加使用氢形成丙酸酯而导致的乙酸酯:丙酸酯比例的降低可能与 CT 抑制CH 4产生有关。但是,在CT浓度与乙酸盐:丙酸酯比例(R 2 = 0.01)或乙酸盐:丙酸酯比例与CH 4之间没有关系。 生产(R 2 = 0.14)。但是,CT和总VFA呈负相关(R 2 = 0.52),而总VFA和CH 4 呈正相关(R 2 = 0.68)。这些建议关系的最简单度量是CT浓度。
因此,CT浓度能否成为 反刍动物肠CH 4减少的令人满意的预测指标?最初的观察结果表明 ,即使在具有相同CT浓度的那些豆类中,每种豆类物种发酵产生的CH 4量也有所不同(Naumann等人,2013b)。然而,线性回归表明,CT和CH 4的 产生呈负相关(R 2 = 0.44)(Naumann等,2013b,c)。仅在相似条件下收集的有限数量的数据集用于测试这种关系。以下用于预测CH 4的非线性指数衰减回归方程 CT的抑制作用是通过增加样本量来开发的:CH 4 = 113.6×Exp(-0.1751×CT)– 2.18;(R 2 = 0.53)(Naumann等人,2015a)。此公式可用于预测潜在的CH浓度4 从含有CT(在CH例如4%CT = 50%的降低的已知浓度底物的发酵产生的4),并进一步用作精密饮食制剂的工具。
研究界有责任开发数学模型,以帮助减轻 反刍动物CH 4的产生和排放,以努力提高动物的生产效率,同时最大程度地减少生产对环境的影响。使用预测模型来估计诸如CT等饲料和饲料成分对瘤胃CH 4 产生和排放的影响是一种方法。单宁浓缩可能会影响反刍动物产甲烷的过程,从而抑制CH 4 排放。问题是:我们如何预测 CT对瘤胃CH 4的抑制作用,并发展对现有方法的未来改进?
局限性和未来方向
可能,推进CT-动物相互作用的知识的最大局限性与CT的结构多样性及其化学测定有关。如上所述,CT是通过一些不同的共价键模式通过连接黄烷-3-醇亚基的子集而组装的。那么,从一小批黄烷-3-醇亚基和一组确定的黄烷间键合中可以衍生出多少个独特的化学实体(异构体)?使用公式(A m ×B n)计算CT低聚物/聚合物中潜在的异构体数(表1))=可能的异构体数量,其中A =化合物中不同的flavan-3-ol亚基类型的数量,m =化合物中的flavan-3-ol亚基的实际数量,B =不同类型的interflavan的数量化合物中存在的键合,n =化合物中存在的黄酮间键的实际数目。据报道,类似的公式可以确定CT结构异构体的数量(Cheynier,2005 ; Nam et al。,2017)。A列中列出了可能由CT产生的不同独特化合物的数量,其中包含两个不同的flavan-3-ol亚基,并且仅两种类型的flavan之间的键合(表1)。牛痘 (蔓越莓)和 高粱的CT 属于本列中化学结构异构体列出的化合物的类别。 牛痘 和 高粱 CT几乎完全由原花青素(儿茶素和表儿茶素)亚基组成。既 高粱 和 越桔 CT具有4-8 B型interflavan联系。 高粱 也可能含有4-6个B型键,而蔓越莓则含有A型黄酮键。来自普通饲料(例如, L。corniculatus,L。pedunculatus 和 O. viciifolia) 的CT包含PC和PD亚基的不同混合物,增加了CT的复杂性(表1,E和F列)。为了增加复杂性,CT结构与某些来源隔离,例如 葡萄 或 薯os (Tian等人,2012)被发现含有衍生的C3羟基作为没食子酸酯。这可以发生多次,并且可以沿着低聚物/聚合物链的任何位置发生。表1中列出的数字 也没有考虑到flavan-3-ol亚基的对映异构体(镜像图像)偶尔会被分离和表征。这两种情况都会大大增加可能的结构异构体的数量,并增加了从植物材料中鉴定和表征CT的挑战。
表1 可能的缩合单宁异构体数目,取决于唯一的flavan-3-ol亚基的数目和存在的flaflavan键类型
| 一个 | 乙 | C | d | 和 | F | |
|---|---|---|---|---|---|---|
| 黄烷-3-醇单元数 | 两个单位两种债券类型 | 两个单位三种债券类型 | 三单元两种债券类型 | 三单元三种债券类型 | 四单元两种债券类型 | 四单元三种债券类型 |
| 2(二聚体) | 8 | 12 | 18 | 27 | 32 | 48 |
| 3(三聚体) | 32 | 72 | 108 | 243 | 256 | 576 |
| 4(四聚体) | 128 | 432 | 648 | 2187 | 2048 | 6,912 |
| 5(五聚体) | 512 | 2592 | 3888 | 19,683 | 16,384 | 82,944 |
| 6(六聚体) | 2048 | 15,552 | 23,328 | 177,147 | 131,072 | 995,328 |
| 7(庚烷) | 8192 | 93,312 | 139,968 | 1,594,323 | 1,048,576 | 11,943,936 |
| 8(八进制) | 32,768 | 559,872 | 839,808 | 14,348,907 | 8,388,608 | 143,327,232 |
| 9(无名) | 131,072 | 3,359,232 | 5,038,848 | 129,140,163 | 67,108,864 | 1,719,926,784 |
| 10(十进位) | 524,288 | 20,155,392 | 30,233,088 | 1,162,261,467 | 536,870,912 | 20,639,121,408 |
另一个局限性是,许多关注CT-动物相互作用的已发表研究都是使用 体外 技术进行的。 体外 技术是CT生物活性的令人满意的筛选工具,并且与体内 方法相比通常更简单,更可控且成本更低 。但是,体外 方法的结果及其与农场现实的相关性存在局限性 。它们不考虑瘤胃液中的微生物更新,通过率和最终产品的去除。缺乏使用体内 CT-动物相互作用的研究 方法在这方面限制了进展。例如,需要进一步评估非线性指数衰减回归方程,以预测 使用活体动物的反刍动物产生的CH 4排放。今天,我们有了一种预测 CT 抑制CH 4的方法。我们需要基于体内 研究的可靠模型 。
短期研究是推进CT-动物相互作用知识的另一个限制。 Odenyo等。(1997) 证明瘤胃微生物如何适应植物的专门代谢产物。 分别由于线虫和微生物种群的适应,驱虫活性或CH 4排放量减少的响应可能无法持续。为数不多长期的 体内 研究进行由供给的 栗 和 栎橡碗 到羊190和85天,分别(Wischer等人,2014)。肠溶CH 4 在整个实验过程中减排没有持续。但是,某些饮食成分的消化率下降。本研究中使用的植物富含HT而不是CT,这可以解释其抗营养作用以及CH 4缺乏持续下降的原因。需要 使用已知的化学和结构特征使用CT而不是HT 进行长期 体内研究,以更好地了解CT与动物的相互作用和相关的动物反应。但是,由于缺乏大量底物的研究人员受到限制,这些底物包含进行长期体内活检所需的足够浓度的生物活性CT 学习。解决此问题的一种方法是使用富含CT的工业副产品,例如葡萄果渣(一种葡萄酒行业副产品)。但是,此类产品的不一致性带来了其他挑战,并且需要在每次使用之前进行CT分析。
当存在其他植物特有的代谢物时,未来的研究也应侧重于CT生物活性。例如,当其他潜在的瘤胃CH 4降低 剂与CT结合使用时,应建立预测方程。CT与其他植物成分之间存在潜在的相互作用,可能会降低CH 4的 产生(Tedeschi等,2011)。)。除CT外,植物还可以产生有机酸,萜烯及其衍生物,生物碱和氰基糖苷。这些代谢物与饮食中的脂肪,纤维和蛋白质因素相结合,在反刍动物的整体营养,抗营养以及CT的功效中起着参与性作用。确定这些不同的植物特定代谢产物和其他饮食成分在瘤胃中如何相互作用很重要。这些作用在动物营养(或抗营养),营养利用,矿物质结合,产甲烷过程和驱虫活性方面是否具有相加,协同或拮抗作用?
浓度,蛋白质/单宁比和pH。国际食品科学与技术杂志47:875-878。[ 友情链接 ]
Afsana,K .; 滋贺 Ishizuka,S.和Hara,H. 2004年。摄入单宁酸对大鼠铁吸收的减少作用,但对锌,铜和锰的吸收没有影响。生物科学,生物技术和生物化学68:584-592。[ 友情链接 ]
Agle,M。AN,Hristov;Zaman,S .; 施耐德角;Ndegwa,P.和Vaddella,VK2010。瘤胃降解蛋白对瘤胃发酵和奶牛粪便氨气损失的影响。乳业科学杂志93:1625-1637。[ 友情链接 ]
Mguer Aguerre;马萨诸塞州Wattiaux; 鲍威尔(JM);Broderick,GA和Arndt,C.,2011年。奶牛日粮中饲草精料比对甲烷,二氧化碳和氨气排放,泌乳性能和粪便排泄的影响。乳业科学杂志94:3081-3093。[ 友情链接 ]
Mguer Aguerre;卡波佐洛(MC);Lencioni,P .;Cabral,C。和Wattiaux,MA,2016年。在2种日粮粗蛋白水平下,栎树栗栗单宁提取物对奶牛的性能,瘤胃发酵和氮分配的影响。乳业科学杂志99:4476-4486。[ 友情链接 ]
Ahnert,S .;美国迪克霍弗;Schulz,F。和Susenbeth,A.,2015年。瘤胃Quebracho单宁提取物注入对小母牛表观营养物质消化率,氮平衡和尿嘌呤衍生物排泄的影响。畜牧科学177:63-70。[ 友情链接 ]
Al-Dobaib,SN2009。不同水平的Quebracho丹宁酸对Najdi绵羊饲喂苜蓿(Medicago sativa) 干草作为唯一饮食的纳吉迪羊的氮利用和生长性能的影响 。动物科学杂志80:532-541。[ 友情链接 ]
Al-Mamary,M。AH,Molham;Abdulwali,AA和Al-Obeidi,A。2001。日粮高粱鞣质对兔消化酶和矿物质吸收的体内作用。营养研究21:1393-1401。[ 友情链接 ]
马萨诸塞州阿隆索-迪亚兹(Alonso-Diaz);JFJ的Torres-Acosta;Sandoval-Castro,CA和Capetillo-Leal,CM2012。来自山羊和绵羊整个唾液的蛋白质的氨基酸分布及其与从热带植物饲料中提取的鞣酸和单宁的相互作用。小反刍动物研究103:69-74。[ 友情链接 ]
RA,安德森;加利福尼亚州布罗德赫斯特;MM,波兰斯基;WF,Schmidt;汗,A,弗拉纳根(Vlanagan),副总裁;Schoene,NW和Graves,DJ,2004年。从肉桂中提取具有胰岛素样生物活性的多酚A型聚合物,并进行了表征。农业和食品化学学报52:65-70。[ 友情链接 ]
M.Andjelkovic;范·坎普(J. De Meulenaer,B .;Depaemelaere,G .;Socaciu,C .;Verloo,M。和Verhe,R。2006。带有邻苯二酚和没食子酰基的酚酸的铁螯合性能。食品化学98:23-31。[ 友情链接 ]
E.Barrau。法布尔,北;Fouraste,I。和Hoste,H.,2005年。皂角苷(Onobrychis viciifolia Scop。)中生物活性化合物对扭体Haemonchus contortus的幼体迁移的影响:单宁和黄酮糖苷的作用。寄生虫学131:531-538。[ 友情链接 ]
贝克尔(PM);PG,维克塞拉;MCR弗兰森;Vos,RCH;Hall,RD和Beekwilder,J. 2014。(+)儿茶素介导的反刍类温室气体甲烷减排的氢沉机制的证据。代谢组学10:179-189。[ 友情链接 ]
R.Bhatta; 萨拉瓦南 Baruah,L。和Prasad,CS,2015年。含单宁的热带树叶的分级水平对体外瘤胃发酵,原生动物和甲烷总产量的影响。应用微生物学杂志118:557-564。[ 友情链接 ]
布雷恩斯;Viveros,A .;哥妮,我。Centeno,C .;SGSáyago-Ayerdy;Arija,I.和Saura-Calixto,F.2008。葡萄渣浓缩物和维生素E对鸡肉中多酚消化率和抗氧化活性的影响。家禽科学87:307-316。[ 友情链接 ]
马萨诸塞州布鲁克斯; RM,哈维;Johnson,NF和Kerley,MS2012。瘤胃中可降解蛋白质的供应会影响连续培养和ste牛生长中的微生物效率。动物科学杂志90:4985-4994。[ 友情链接 ]
M. Brune;Rossander,L.和Hallberg,L.1989。铁吸收和酚类化合物:不同酚类结构的重要性。欧洲临床营养杂志43:547-558。[ 友情链接 ]
上午,布鲁诺-苏亚雷斯;阿拉巴马州Soares-Pereira;Matos,TJS和Ricardo-da-Silva,JM2011。对 小反刍动物中 羽扇豆(Lupinus angustifolius)种子蛋白级分的体外肠道消化率,葡萄(Vitis vinifera)种子浓缩单宁 的影响的初步结果 。动物生理与动物营养学杂志95:456-460。[ 友情链接 ]
Buddle,BM;丹尼斯(Denis)GT Attwood;E.Altermann,E。詹森,PH;Ronimus,RS;皮纳雷斯-帕蒂诺(CS);Muetzel,S.和Neil Wedlock,D.2011。减少在牧场上放牧的反刍动物甲烷排放的策略。兽医杂志188:11-17。[ 友情链接 ]
Buzzini,P.,Arapitsas,P .; M.Goretti;E.Branda;B.图尔切蒂 Pinelli,P .;Ieri,F.和Romani,A.2008。可水解单宁的抗微生物和抗病毒活性。药物化学迷你评论8:1179-1187。[ 友情链接 ]
萨尔瓦多州 雪貂,A。雷诺兹(CK);NB的Kristensen和AM AM的van Vuuren,2010年。反刍动物对氮的优化利用策略。动物4:1184-1196。[ 友情链接 ]
佳能F. Ballivian,R .; Chirot,F .; Antoine,R .;Sarni-Manchado,体育;Lemoine,J.和Dugourd,P.2011。与单宁结合后唾液内在无序蛋白的折叠。美国化学学会杂志133:7847-7852。[ 友情链接 ]
MG卡帕伊;沃尔夫 Pinna,W.和Kamphues,J.2013。猪使用内源性脯氨酸来应对橡子(Quercus pubescens Willd。)混合日粮中富含可 水解单宁的饮食。牲畜科学155:316-322。[ 友情链接 ]
Carnovale,V .; Britten,M。Couillard,C.和Bazinet,L.2015。钙对表没食子儿茶素-3-没食子酸酯和β-乳球蛋白之间相互作用的影响。食品研究国际杂志77:565-571。[ 友情链接 ]
Chamorro,S。Viveros,A .;Centeno,C .;罗梅罗角; Arija,I.和Brenes,A.2013。饮食中的葡萄籽提取物对肉仔鸡生长性能,氨基酸消化率,血浆脂质和矿物质含量的影响。动物7:555-561。[ 友情链接 ]
AJ查尔顿;新泽西州巴克斯特;汗,ML;Moir,AJG;E.哈斯兰;Davies,AP和Williamson,MP2002。多酚/肽结合和沉淀。农业和食品化学学报50:1593-1601。[ 友情链接 ]
Cheynier,V.2005。食品中的多酚比通常认为的要复杂。美国临床营养学会81:223S-229S。[ 友情链接 ]
RG,克里斯滕森;杨SY; Eun,J.-S .;杨,AJ;Hall,JO和MacAdam,JW,2015年。饲喂鸡足三叶草干草对中性洗涤剂纤维消化,氮利用效率和奶牛泌乳性能的影响。乳业科学杂志98:7982-7992。[ 友情链接 ]
Coblentz,WK和Grabber,JH,2013年。由于单宁浓度升高,苜蓿和鸟足三叶草干草和青贮饲料的原位蛋白质降解。乳业科学杂志96:3120-3137。[ 友情链接 ]
de Freitas,V.和Mateus,N.2002。唾液蛋白单宁酸聚集物的比浊法研究。粮食与农业科学学报82:113-119。[ 友情链接 ]
急诊部迪维尔;Givens,DI和Mueller-Harvey,I.,2010。栗子和含羞草单宁青贮饲料:对绵羊的影响因表观消化率,氮素利用和损失而有所不同。动物饲料科学与技术157:129-138。[ 友情链接 ]
Delehanty,JB;BJ约翰逊;TE的Hickey;Pons,T。和Ligler,FS2007。植物原花青素对脂多糖的结合和中和作用。天然产物杂志70:1718-1724。[ 友情链接 ]
O. Desrues;Fryganas,C .;HM Ropiak; 我是Mueller-Harvey;Enemark,HL和Thamsborg,SM2016。黄烷醇单体和缩合单宁的化学结构对体外对牛线虫驱虫活性的影响。寄生虫学143:444-454。[ 友情链接 ]
美国迪克霍弗;Ahnert,S。和Susenbeth,A。2016。quebracho单宁提取物对瘤胃发酵以及小母牛微生物质量的产量和组成的影响。动物科学杂志94:1561-1575。[ 友情链接 ]
蒙大拿州迪特曼;美国龙格;朗(RA)Moser D .; Galeffi,C .; Kreuzer,M.和Clauss,M.2014。骆驼科动物的甲烷排放。PLoS 1 9:1-9。[ 友情链接 ]
马萨诸塞州多布雷瓦; 弗雷泽(RA)我是Mueller-Harvey;洛杉矶克利夫顿; Gea,A.和Green,RJ,2011年。戊铝酰葡萄糖与两种球蛋白的结合是通过多个表面位点发生的。生物大分子12:710-715。[ 友情链接 ]
多西,RR;贝伦格(A.)PG,托拉尔;Hervás,G。和Frutos,P.,2013年。施用栎栎幼叶对瘤胃发酵的影响,与牛的橡木单宁中毒有关。动物生理与动物营养杂志97:48-57。[ 友情链接 ]
CM Dschaak;威廉姆斯,CM;霍尔特(MS)Eun,J.-S .;Young,AJ和Min,BR,2011年。补充浓缩单宁提取物对泌乳奶牛的摄入,消化,瘤胃发酵和产奶的影响。乳业科学杂志94:2508-2519。[ 友情链接 ]
Elrod,CC和Butler,WR,1993。饲喂过量瘤胃可降解蛋白质的小母牛的繁殖力降低和子宫pH值改变。动物科学杂志71:694-701。[ 友情链接 ]
蒙大拿州Engström; 卡罗宁(M. JR Ahern;北贝特;佩雷(B.)Hoste先生,H.和Salminen,植物水解单的JP 2016年化学结构揭示针对的卵孵化和活力它们的体外活性 捻转血矛线虫 线虫。农业与食品化学学报64:840-851。[ 友情链接 ]
S.Fairweather-Tait; Z.Piper;Fatemi,SJA和Moore,GR1991。茶对大鼠铁和铝代谢的影响。英国营养杂志65:61-68。[ 友情链接 ]
新界Faithfull,1984年。饲料的体外消化率-发酵的一个世纪。粮食与农业科学学报。35:819-826。[ 友情链接 ]
Feng,WY2006。《绿茶儿茶素的代谢:概述》。当前药物代谢7:755-809。[ 友情链接 ]
菲利皮希(LJ);朱建。Oelrichs,P .;蒙大拿州阿尔萨拉米;Doig,AJ;Cao,GR和English,PB,1991。《长叶榄仁的肝毒性和肾毒性原理》。兽医学研究50:170-177。[ 友情链接 ]
富,LY;陆Y; Howell,AB和Vorsa,N。2000。蔓越莓中的A型原花青素三聚体,可抑制尿毒症性p纤维化大肠杆菌的附着 。 天然产物杂志63:1225-1228。[ 友情链接 ]
格塔切夫;皮特罗夫(W. DH普特南;丹德卡(A.)Goyal,S.和Depeters,EJ2008。向苜蓿干草中添加没食子酸,单宁酸或quebracho单宁对体外瘤胃发酵和微生物蛋白质合成的影响。动物饲料科学与技术140:444-461。[ 友情链接 ]
Glahn,RP和Wortley,GM,2002年。《植酸,单宁酸和ZnCl2抑制铁吸收:使用体外消化/ Caco-2细胞模型进行的研究。农业和食品化学学报50:390-395。[ 友情链接 ]
贡萨尔维斯河;Mateus,N.和De Freitas,V.2011。缩合的单宁酸抑制α-淀粉酶活性。食品化学125:665-672。[ 友情链接 ]
Grainger,C.和Beauchemin,KA2011。能否在不降低反刍动物产量的情况下降低反刍动物的肠甲烷排放量?动物饲料科学与技术166-167:308-320。[ 友情链接 ]
Grainger,C .;克拉克,T。MJ奥尔德斯;KA Beauchemin; McGinn,SM;Waghorn,GC和Eckard,RJ,2009年。潜在使用的金合欢(Acacia mearnsii)浓缩单宁酸可以减少放牧奶牛的甲烷排放和氮排泄。加拿大动物科学杂志89:241-251。[ 友情链接 ]
Greger,JL和Lyle,BJ 1988年。喂养各种水平和类型的茶的大鼠的铁,铜和锌代谢。营养杂志118:52-60。[ 友情链接 ]
Hagerman,AE和Butler,LG 1978年。蛋白质沉淀法,用于单宁的定量测定。农业与食品化学学报26:809-812。[ 友情链接 ]
Hagerman,AE和Butler,LG 1981年。原花色素蛋白相互作用的特异性。生物化学杂志256:4494-4497。[ 友情链接 ]
AE哈格曼;康涅狄格州罗宾斯;Y.Weerasuriya;Wilson,TC和McArthur,C.1992。与消化有关的单宁化学。Journal of Range Management 45:57-62。[ 友情链接 ]
Hagerman,AE和Robbins,CT,1993。单宁结合唾液蛋白相对于哺乳动物饮食选择的特异性。加拿大动物学杂志71:628-633。[ 友情链接 ]
AE哈格曼;Rice,ME和Ritchard,NT,1998。两种单宁,五烷基戊糖和epicatechin16(4-8)儿茶素(原花青素)的蛋白质沉淀机理。农业和食品化学学报46:2590-2595。[ 友情链接 ]
Hagerman,AE,2011年。单宁手册。俄亥俄州牛津的迈阿密大学。可在以下网站上找到:< http://www.users.muohio.edu/hagermae/ >。访问日期:2016年6月27日。[ 链接 ]
Hagerman,AE,2012年。《多酚-蛋白质复合物的五十年》。第71-97页。于:多酚研究的最新进展。卷 3.第三版。Cheynier,V。Sarni-Manchado,P。和Quideau编辑。英国牛津的John Wiley&Sons,Ltd.。[ 友情链接 ]
哈林,DA;Scharenberg,A .; Heckendorn,F .;多米(F.)A.Luscher;Maurer,V .; Suter,D.和Hertzberg,H.2008。单宁草料植物:对绵羊寄生线虫的农艺性能,适口性和功效。可再生农业和粮食系统23:19-29。[ 友情链接 ]
爱荷华州哈桑;EA的Elzubeir和AH,2003年的El Tinay。饲喂低或高单宁含量日粮的肉鸡的矿物质生长和表观吸收。热带动物健康与生产35:189-196。[ 友情链接 ]
Hoste,H。Martinez-Ortiz-De-Montellano,C .;F.Manolaraki;布鲁内特,S。Ojeda-Robertos,N。I. Fourquaux;Torres-Acosta,JFJ和Sandoval-Castro,CA2012。生物活性富含单宁的热带和温带豆科植物对线虫感染的直接和间接影响。兽医寄生虫学186:18-27。[ 友情链接 ]
加州大学Hymes-Fecht; 乔治亚州布罗德里克;Muck,RE和Grabber,JH2013。用鸟足三叶青贮代替混合饲料中的紫花苜蓿或红三叶草青贮可提高泌乳奶牛的产量。乳业科学杂志96:460-469。[ 友情链接 ]
Jansman,AJM,1993年。简单气孔动物饲料中的单宁酸。营养研究评论6:209-236。[ 友情链接 ]
Jayanegara,A .;Goel,G .;Makkar,HPS和Becker,K.,2015年。纯化的可水解和缩合单宁对甲烷排放,瘤胃发酵和体外微生物种群的影响之间的差异。动物饲料科学与技术209:60-68。[ 友情链接 ]
Jin L. 王Y;公元Iwaasa;徐中 席伦贝格,国会议员;张永刚; Liu,XL和McAllister,TA,2012年。单宁浓缩对 两个生长阶段收获的紫色三叶草(Dalea purpurea Vent。)瘤胃降解性的影响 。动物饲料科学与技术176:17-25。[ 友情链接 ]
约翰逊(NF);利斯,我;Kerley,MS和Naumann,HD,2015年。单宁含豆类饲料对体外粗蛋白降解的影响。动物科学杂志93(增刊2):97。[ 友情链接 ]
Kariuki,I.和Norton,B. 2008年。绵羊胃肠道中浓缩的单宁结合的饮食蛋白质的消化。动物饲料科学与技术142:197-209。[ 友情链接 ]
Khokhar,S.和Owusu-Apenten,RK2003。酚类化合物的铁结合特征:一些初步的构效关系。食品化学81:133-140。[ 友情链接 ]
Klongsiriwet,C .;J. 阿联酋威廉姆斯;我是Mueller-Harvey;威廉姆森(Williamson),新兴市场公司(EM)和赫斯特(Hoste)H.,2015年。类黄酮单体和缩合的单宁酸协同抑制捻转金眼梅的出壳。国际寄生虫学药物和耐药性杂志5:127-134。[ 友情链接 ]
RA,Kohn; Dinneen,MM和Russek-Cohen,E.2005。使用血尿素氮预测牛,羊,山羊,马,猪和大鼠的氮排泄和氮利用效率。动物科学杂志83:879-889。[ 友情链接 ]
熊本市 Sonda,T .; Nagayama,K.和Tabata,M.2001。pH和金属离子对儿茶素抗氧化活性的影响。生物科学,生物技术和生物化学65:126-132。[ 友情链接 ]
库玛(Kumar)LC,乔德哈里;Agarwal,N.和Kamra,DN2014。饲喂单宁降解细菌培养物(解链球菌TDGB 406)对饲喂半棕榈栎叶的山羊的养分利用率,尿嘌呤衍生物和生长性能的影响。动物生理学和动物营养学杂志98:879-885。[ 友情链接 ]
SR拉文;Chen,Z.和Abrams,SA,2010年。单宁酸对稻草色果蝠(Eidolon helvum)中铁吸收的影响 。 动物园生物学29:335-343。[ 友情链接 ]
Lavin,SR2012。植物酚类及其在减轻野生动物铁过载中的潜在作用。动物园野生动物医学杂志43:S74-S82。[ 友情链接 ]
Le Bourvellec,C。和Renard,CMGC,2012年。多酚与大分子之间的相互作用:定量方法和机理。《食品科学与营养评论》 52:213-248。[ 友情链接 ]
DM,Leathwick和RB,Besier,2014年。《大洋洲战略和经验》中放牧反刍动物的驱虫抗药性管理。兽医寄生虫学204:44-54。[ 友情链接 ]
李SH;PL Shinde;崔俊杰 IK,Kwon;李建国 朴,SI;Cho,WT和Chae,BJ,2010。补充鞣酸对断奶猪生长性能,血液血液学,铁状态和粪便菌群的影响。家畜科学131:281-286。[ 友情链接 ]
李志平; 刘HL; 李根元 鲍克; 王K 徐成 杨YF;Yang,FH和Wright,A.-DG,2013年。富含丹宁酸和高纤维饲料的瘤胃细菌群落的分子多样性,摄食 于中国的国产梅花鹿 (Cervus nippon)。BMC微生物学13:1-12。[ 友情链接 ]
MM Lorenz;阿尔卡法吉(L. E.Stringano;Nilsson,S .; Mueller- Harvey,I.和Udén,P.2014。单宁缩合结构与其在瘤胃中沉淀饲料蛋白质的能力之间的关系。粮食与农业科学学报94:963-968。[ 友情链接 ]
Lou,H .;Y.山崎 佐佐木 内田,M .; Tanaka,H.和Oka,S.1999。来自花生皮的A型原花色素。植物化学51:297-308。[ 友情链接 ]
马哈茂德,S。阿里(H.)Ahmad,F.和Iqbal,Z.2014。不同高粱品种中单宁的估计及其对养成的白色来格霍恩层中养分消化率和某些矿物质吸收的影响。国际农业与生物学报16:217-221。[ 友情链接 ]
梅特兰 Nierenstein,M .;加利福尼亚州米切尔;GR,林奇;诺曼(美国);休斯(EB)休斯和巴格诺尔(Bagnall),HH1936。茶和咖啡,特别提及其生物碱和单宁酸。分析师61:288-314。[ 友情链接 ]
曼吉诺,J。Peterson,K.和Jacobs,H.2003。开发一种排放模型来估算牛肠发酵产生的甲烷。美国环境。Prot。机构。可在以下网址获得:< http://www.epa.gov/ttnchie1/conference/ei12/green/mangino.pdf >。访问日期:2016年1月14日。[ 链接 ]
麦克阿瑟角;Sanson,G。和Beal,AM,1995年。哺乳动物唾液中富含脯氨酸的蛋白质:在口服体内稳态中的作用和抵消膳食单宁的作用。化学生态杂志21:663-691。[ 友情链接 ]
麦当劳(McDonald);Mila,I.和Scalbert,A.1996。植物多酚对金属离子的沉淀:最佳条件和沉淀来源。农业与食品化学学报44:599-606。[ 友情链接 ]
WC McNabb;瓦格霍恩(Waghorn)Peters,JS和Barry,TN,1996年。莲花梗中单宁的浓缩对瘤胃和Rubisco消化位点中1,5-双磷酸核糖1,5-二磷酸羧化酶蛋白的溶解和降解的影响。英国营养杂志76:535-549。[ 友情链接 ]
WC McNabb;彼得斯,JS;富,LY;Waghorn,GC; Jackson,FS,1998。从几种草料制备的缩合单宁对1,5-双磷酸核糖羧化酶(Rubisco)蛋白的体外沉淀以及胰蛋白酶(EC 2.4.21.4)和糜蛋白酶(EC)的消化作用。 2.4.21.1)。粮食与农业科学学报77:201-212。[ 友情链接 ]
CS McSweeney;Collins,EMC;Blackall,LL和Seawright,AA2008。抗营养因子综述限制了相思树作为反刍动物饲料的潜在用途。动物饲料科学技术147:158-171。[ 友情链接 ]
NR的Merchen和EC的Titgemeyer,1992。处理不断增长的反刍动物的氨基酸供应。动物科学杂志70:3238-3247。[ 友情链接 ]
敏,BR;田纳西州巴里; Attwood,GT和McNabb,WC2003。浓缩单宁对反刍动物饲喂新鲜温带草料的营养和健康的影响:综述。动物饲料科学与技术106:3-19。[ 友情链接 ]
敏,BR;我们是Pinchak;Fulford,JD和Puchala,R.2005。《小麦草场膨胀动态,体外瘤胃气体产生以及缩合单宁酸的潜在膨胀缓解作用。动物科学杂志83:1322-1331。[ 友情链接 ]
敏,BR;S·索莱曼 特里尔(Terrill);Ramsay,A.和Mueller-Harvey,I.2015。含单宁的松树皮地面饮食对肉山羊营养物质消化,氮平衡和矿物质balance留的影响。动物科学与生物技术杂志6:25。[ 友情链接 ]
美联社Minho;ICS,Bueno;Louvandini,H。杰克逊(F. Gennari,SM和Abdalla,AL,2008年。相思木单宁提取物对绵羊胃肠道寄生虫的控制作用。动物饲料科学与技术147:172-181。[ 友情链接 ]
Mladenka,P .; Macakova,K .;Filipsky,T .;Zatloukalova,L .; Jahodár,L .; Bovicelli,P .;Silvestri,IP;Hrdina,R.和Saso,L.2011。类黄酮的铁螯合活性的体外分析。无机生物化学杂志105:693-701。[ 友情链接 ]
痣,S。Butler,LG和Iason,G.,1990年。《草食动物对膳食单宁的防御:对哺乳动物脯氨酸丰富的唾液蛋白的调查。生化系统学与生态学18:287-293。[ 友情链接 ]
莫纳加斯(M. Urpi-Sarda,M .;桑切斯-帕坦(F. 罗洛拉河; 加里多,我。戈麦斯-科尔多瓦(C. Andres-Lacueva,C。和Bartolomé,B。2010。《饮食性flavan-3-ols的代谢和微生物生物转化及其代谢物的生物活性的见解》。食品与功能1:233-253。[ 友情链接 ]
Mueller-Harvey,I.2006。揭示单宁酸在动物营养和健康中的难题。粮食与农业科学学报86:2010-2037。[ 友情链接 ]
Nam,J.-W .; RS Phansalkar;华盛顿特区 JB的McAlpine;莱姆·克劳斯(AA);威达(CMP);Gan,L.-S .; Bedran-Russo,A .;陈信义 和Pauli,GF2017。具有牙本质生物修饰作用的天然寡聚原花色素的绝对构型。有机化学杂志82:1316-1329。[ 友情链接 ]
瑙曼,高清;缪尔(JP);BD兰伯特;密苏里州特德斯基(Tedeschi)和MM柯思曼(MM)2013a 反刍动物环境中的单宁浓缩:生物活性的观点。农业科学学报1:8-20。[ 友情链接 ]
瑙曼,高清;路易斯安那州Tedeschi;缪尔(JP);BD的Lambert和MM的科斯曼2013b。暖季多年生豆科植物缩合单宁的分子量对体外瘤胃甲烷产生的影响。生化系统学与生态学50:154-162。[ 友情链接 ]
瑙曼,高清;路易斯安那州Tedeschi;AE哈格曼;Lambert BD和Muir,JP 2013c。暖季多年生豆类植物单宁缩合的甲烷排放和蛋白质沉淀能力。第491-492页。在:可持续动物生产中的能量和蛋白质代谢与营养,EAAP出版物第134号。E. Kebreab和H. Lapierre编辑。瓦赫宁根学术出版社。荷兰瓦赫宁根。[ 友情链接 ]
瑙曼,高清;SA,阿姆斯特朗;BD兰伯特;缪尔(JP);密苏里州特德斯基(Tedeschi)和MM柯斯曼(MM)2014a。豆科植物缩合单宁的分子量和浓度对捻转血矛线虫体外幼虫迁移抑制的影响。兽医寄生虫学199:93-98。[ 友情链接 ]
瑙曼,高清;AE哈格曼;BD兰伯特;缪尔(JP);密苏里州特德斯基(Tedeschi)和MM柯斯曼(Kothmann),2014b。暖季多年生豆科植物中单宁缩合的分子量和蛋白质沉淀能力。植物相互作用杂志9:212-219。[ 友情链接 ]
瑙曼,高清;新墨西哥州切里; 路易斯安那州Tedeschi;缪尔(JP)和兰伯特(BD 2014c)。反刍动物中蛋白质可沉淀的多酚(缩合单宁)对蛋白质结合和蛋白质消化的概念模型。动物科学杂志92(E-增刊2):910。[ 友情链接 ]
瑙曼,高清;马萨诸塞州丰塞卡(Fonseca)和LO特德斯基(Tedeschi)2015a。使用非线性指数衰减回归分析预测缩合单宁对瘤胃甲烷的抑制作用。动物科学杂志93:5341-5345。[ 友情链接 ]
瑙曼,高清;BD兰伯特;SA,阿姆斯特朗;马萨诸塞州丰塞卡; 路易斯安那州Tedeschi;JP Muir和Ellersieck,MR 2015b。在基于玉米苜蓿的基质中,用圆锥花序的三叶草或绢丝桃代替紫花苜蓿对体外瘤胃甲烷产生的影响。乳业科学杂志98:3980-3987。[ 友情链接 ]
Ng,F。基特尔曼,S。帕切特(ML);GT Attwood;詹森,PH;Rakonjac,J。和Gagic,D.,2016年。利用瘤胃产甲烷菌甲烷甲烷短杆菌反刍动物M1产生的氢结合素结合了广泛的产氢微生物。环境微生物学18:3010-3021。doi:10.1111 / 1462-2920.13155。[ 友情链接 ]
O’Donovan,L.和Brooker,JD2001。可水解和缩合单宁对解脂链球菌(S. caprinus) 和牛链球菌的生长,形态和代谢的影响 。微生物学147:1025-1033。[ 友情链接 ]
Aden的Odenyo;Osuji,PO;Karanfil,O.和Adinew,K.1997。金合欢作为绵羊蛋白质补充的微生物学评估。动物饲料科学与技术65:99-112。[ 友情链接 ]
奥兰迪(美国);Goz Kozloski;阿尔维斯(TP);密苏里州梅斯基塔(Mesquita)和南卡罗莱纳州阿维拉(2015)。饲草饲草和浓缩物中含有不断增加的金合欢鞣 质单宁提取物的牛的消化率,瘤胃发酵和十二指肠氨基酸通量 。动物饲料科学与技术210:37-45。[ 友情链接 ]
奥兹达尔(T. Capanoglu,E.和Altay,F.2013。蛋白质-酚相互作用和相关变化的综述。国际食品研究杂志51:954-970。[ 友情链接 ]
Patra,AK和Saxena,J.2011。利用膳食单宁改善瘤胃代谢和反刍动物营养。粮食与农业科学学报91:24-37。[ 友情链接 ]
佩雷斯·格雷戈里奥(MR)Mateus,N.和De Freitas,V.2014。通过质谱法(MALDI-TOF-TOF和FIA-ESI-MS)快速筛选和鉴定唾液中新的可溶性单宁唾液蛋白聚集体。Langmuir 30:8528-8537。[ 友情链接 ]
Perron,NR和Brumaghim,JL2009。多酚化合物与铁结合的抗氧化机理综述。细胞生物化学与生物物理学53:75-100。[ 友情链接 ]
Poncet-Legrand,C .;埃德尔曼(A.)J.-L. Putaux;卡塔拉德(D. Sarni-Manchado,P.和Vernhet,A.2006。与flavan-3-ols单元的聚脯氨酸相互作用:分子结构和多酚/蛋白质比率的影响。食品水胶体20:687-697。[ 友情链接 ]
鲍威尔(JM);密西根州阿圭尔(Aguerre)和马萨诸塞州瓦蒂(Wattiaux),2010年。单宁可从模拟的奶牛场地板中提取减少的氨气排放。环境质量杂志40:907-914。[ 友情链接 ]
Quideau,S。德菲,D .;Douat-Casassus,C.和Pouységu,L.2011。植物多酚:化学性质,生物活性和合成。Angewandte Chemie国际版50:586-621。[ 友情链接 ]
J. Fryganas,C .;HM Ropiak; 拉姆齐(A. Mueller-Harvey,I。和Hoste,H.,2015年。小型反刍动物对弯曲变形杆菌(Haemonchus contortus)或粘虫(Trichostrongylus colubriformis)的驱虫活性受缩合单宁结构特征的影响。农业与食品化学学报63:6346-6354。[ 友情链接 ]
Ramin,M.和Huhtanen,P.2013。预测反刍动物甲烷排放量的方程式的开发。乳业科学杂志96:2476-2493。[ 友情链接 ]
LM雷东多;宾夕法尼亚州查卡纳; Dominguez,JE和Fernandez Miyakawa,ME,2014。使用单宁替代家禽中抗菌生长促进因子的观点。微生物学前沿5:1-7。[ 友情链接 ]
Roche,JF,2006年。《奶牛营养管理对生殖效率的影响》。动物生殖科学96:282-296。[ 友情链接 ]
萨米纳森(Saminathan)Tan,HY;CC,Sieo;N.阿卜杜拉;黄,CMVL;Abdulmalek,E。和Ho,YW,2014年。来自白斑杂种的缩合单宁馏分的聚合度,分子量和蛋白质结合亲和力。分子19:7990-8010。[ 友情链接 ]
Santos-Buelga,C.和Scalbert,A.2000。原花青素和单宁类化合物–性质,发生率,饮食摄入量以及对营养和健康的影响。粮食与农业科学学报80:1094-1117。[ 友情链接 ]
A.Scalbert 莫兰德;Manach,C。和Rémésy,C。2002。肠道中多酚的吸收和代谢及其对健康的影响。生物医学和药物疗法56:276-282。[ 友情链接 ]
Schwab,CG,1995。反刍动物的保护蛋白和氨基酸。第115-141页。在:动物饲料和动物饲料中的生物技术。华莱士,RJ和切森,A。编。VCH,纽约,纽约。[ 友情链接 ]
出售,DR;里德(WM);Chrisman,CL和Rogler,JC1984。受高粱单宁影响的粘蛋白排泄和肠道形态。营养报告国际31:1369-1374。[ 友情链接 ]
SA Shaik;泰里尔(Terrill);JE,Miller;B.Kouakou;甘南(Kannan);卡普兰(Kaplan)Burke,JM和Mosjidis,JA,2006年。Sericea lespedeza干草是天然的驱虫剂,可防止山羊胃肠道线虫感染。兽医寄生虫学139:150-157。[ 友情链接 ]
AH,史密斯;Zoetendal,E。和Mackie,RI,2005年。克服膳食单宁抑制作用的细菌机制。微生物生态学50:197-205。[ 友情链接 ]
Soares,S .;维托里诺河; 奥索里奥(H. 费尔南德斯(A. Venâncio,A。马特乌斯,北;Amado,F.和De Freitas,V.2011。人类唾液蛋白家族对食物多酚的反应性。农业与食品化学学报59:5535-5547。[ 友情链接 ]
Soares,S .;N.Mateus和V.De Freitas,2012a。碳水化合物通过单宁浓缩抑制唾液蛋白沉淀。农业与食品化学学报60:3966-3972。[ 友情链接 ]
Soares,S .;苏萨(A.);N.Mateus和V.De Freitas,2012b。单宁浓缩对红酒涩味的影响。化学感官37:191-198。[ 友情链接 ]
斯潘塞(Spencer),厘米;蔡Y;马丁·R。上海加夫尼;PN,古尔德;Magnolato,D .;Lilley,TH和Haslam,E.,1988年。《多酚络合-一些想法和观察》。植物化学27:2397-2409。[ 友情链接 ]
Tamilmani,P.和Pandey,MC,2016年。多酚的铁结合效率:抗坏血酸和乙二胺四乙酸对儿茶酚和没食子酰基的影响比较。食品化学197:1275-1279。[ 友情链接 ]
路易斯安那州Tedeschi;TR,卡拉威;Muir,JP和Anderson,RC,2011。饲料添加剂的潜在环境效益和其他反刍动物生产策略。Revista Brasileira de Zootecnia 40:291-309。[ 友情链接 ]
路易斯安那州Tedeschi;Ramírez-Restrepo,CA和Muir,JP,2014年。开发一种概念化的模型,该模型探讨了单宁浓缩对反刍动物生产的益处。动物8:1095-1105。[ 友情链接 ]
泰里尔(Terrill);JA,Mosjidis;特拉华州摩尔;SA Shaik;JE,Miller;伯克(JM);Muir,JP和Wolfe,R.,2007年。制粒对Sericea lespedeza干草作为山羊天然驱虫剂功效的影响。pdf。兽医寄生虫学146:117-122。[ 友情链接 ]
特里尔(Terrill);戴克斯(Dykes)Shaik,S .;米勒,J。B.Kouakou;甘南(Kannan);Burke,J.和Mosjidis,J.2009。绢丝藻干草作为山羊的天然驱虫药的功效:剂量滴定研究。兽医寄生虫学163:52-56。[ 友情链接 ]
K.Theodoridou;奥弗雷尔(J. D.Andueza;Pourrat,J .;Le Morvan,A .;E.Stringano;Mueller-Harvey,I.和Baumont,R.2010。新鲜皂素(Onobrychis viciifolia)中的单宁浓缩 对绵羊体内和原位消化的影响。动物饲料科学技术160:23-38。[ 友情链接 ]
田Y;邹乙;Li C.-M .;杨建旭,S.-F. 和Hagerman,AE,2012年。高分子量柿子单宁是体内和体外的有效抗氧化剂。国际食品研究45:26-30。[ 友情链接 ]
Tshuma,T .; 霍姆(德国);Fosgate,GT和Lourens,DC,2014年。Bonsmara小母牛在不同管理系统内的预培养血尿素氮浓度和生殖性能。热带动物健康与生产46:1023-1030。[ 友情链接 ]
美国环保局。2016。美国温室气体清单报告:1990-2014。可在以下网址获取:< https://www3.epa.gov/climatechange/ghgemissions/usinventoryreport.html >。访问日期:2016年7月1日。[ 链接 ]
Van Duinkerken,G .;安德烈(G.)Smits,MC和Monteny,GJ和Sebek,LB,2005年。瘤胃可降解蛋白平衡和饲草类型对散装牛奶尿素浓度和氨气排放的影响。乳业科学杂志88:1099-1112。[ 友情链接 ]
瓦尔加斯·麦加纳(J. Aguilar-Caballero,AJ;JFJ的Torres-Acosta;加利福尼亚州桑多瓦尔-卡斯特罗;Hoste,H.和Capetillo-Leal,CM2013。热带单宁含量丰富的饲料摄入会改变生长绵羊的唾液结合能力。动物7:1921-1924。[ 友情链接 ]
瓦格霍恩(Waghorn)密歇根州乌利亚特;John,A。和Fisher,MT,1987年。浓缩单宁对以莲花角corn为食的绵羊的氨基酸和其他营养物质的消化位点的 影响。英国营养杂志57:115-126。[ 友情链接 ]
瓦格霍恩(Waghorn)内华达州谢尔顿(Shelton)和WCN McNabb(1994年)。莲中单宁的浓缩对其绵羊营养价值的影响。1.非氮方面。农业科学杂志123:99-107。[ 友情链接 ]
瓦格霍恩(Waghorn)爱达荷州谢尔顿;WC McNabb和SN 1994b的McCutcheon。莲花梗中单宁的浓缩对其绵羊营养价值的影响。2.氮方面。农业科学杂志123:109-119。[ 友情链接 ]
Waghorn,G.,2008年。膳食缩合单宁对可持续绵羊和山羊生产的有益和有害影响-进展和挑战。动物饲料科学与技术147:116-139。[ 友情链接 ]
康涅狄格州韦斯特伍德; IJ精益;Garvin,JK和Wynn,PC,2000年。遗传价值和日粮蛋白质降解性对泌乳奶牛的影响。乳业科学杂志83:2926-2940。[ 友情链接 ]
威廉姆斯,CM;Eun,J.-S .;CM Dschaak;麦克亚当,JW;Min,BR和Young,AJ2010。案例研究: 连续培养中的鸡足三叶干草(Lotus corniculatus L.)干草的体外瘤胃发酵特性 。专业动物科学家26:570-576。[ 友情链接 ]
威廉姆斯,CM;Eun,J.-S .;麦克亚当,JW;杨,AJ;Fellner,V.和Min,BR,2011年。在连续混合瘤胃微生物的连续培养中,含有缩合单宁的豆类饲料对甲烷和氨产生的影响。动物饲料科学与技术166-167:364-372。[ 友情链接 ]
Wischer,G。Greiling,上午;Boguhn,J。Steingass,H。Schollenberger,M。Hartung,K.和Rodehutscord,M.2014。长期补充栗子和valonea提取物对绵羊甲烷释放,消化率和氮排泄的影响。动物8:938-948。[ 友情链接 ]
伍德,C .;方SG 亨特 Streich,WJ和Clauss,M.,2003年。《狐猴中铁的吸收增加:定量筛查和饮食预防评估》。美国灵长类动物杂志61:101-110。[ 友情链接 ]
ren,AF;Cleary,M。弗朗茨C. Melton,S.和Norris,L.2013。一种大鼠葡萄籽提取物(IH636)的90天口服毒性研究。农业与食品化学学报50:2180-2192。[ 友情链接 ]
Yun,S .; Habicht,J.-P .;Miller,DD和Glahn,RP2004。体外消化/ Caco-2细胞培养系统可准确预测抗坏血酸和多酚化合物对人体铁生物利用度的影响。营养杂志134:2717-2721。[ 友情链接 ]
收到:2017年6月8日; 接受:2017年9月5日
* 通讯作者:naumannhd@missouri.edu
这是根据知识共享署名许可协议条款分发的开放访问文章,该文章允许在适当引用原始作品的情况下,在任何介质中进行不受限制的使用,分发和复制。