
单宁酸替代饲用抗生素用于农场动物生产的潜力和挑战
摘要
天然存在的植物化合物(包括 单宁, 皂苷 和 精油) 被广泛评估为饲料中抗生素的天然替代品。单宁酸是一组多酚化合物,广泛存在于植物区域,并具有多种生物活性,包括抗微生物,抗寄生虫,抗病毒,抗氧化剂,抗炎,免疫调节等。因此,单宁酸是开发中的主要研究对象。饲料中抗生素的天然替代品。强大的蛋白质亲和力是植物单宁的公认特性,已成功应用于 反刍动物 营养以减少瘤胃中蛋白质的降解 ,从而提高蛋白质利用率和动物生产效率。 反刍动物日粮中含有单宁 草料以控制动物牧场膨胀,肠道寄生虫和致病菌负荷是反刍动物中单宁的另外三个重要应用。单宁传统上一直被认为是单胃 动物和家禽的“抗营养因子”,但是最近的研究表明,其中的一些单宁 以适当的方式施用可以改善肠道微生物生态系统,增强肠道健康,从而提高生产性能。植物单宁酸作为饲料中抗生素的替代品的适用性取决于许多因素,这些因素导致其观察到的功效差异很大。
关键词
单宁 生物活动 饲料中的抗生素 天然饲料添加剂 农场动物
1.简介
在农场动物生产中促进生长的抗生素已经使用了几十年,并被证明可以有效提高动物和 家禽的生产 效率。但是,普遍认为使用抗生素作为生长促进剂可促进家畜中抗生素抗性微生物的进化和/或选择(Chattopadhyay,2014年)。在过去的几十年中,已经进行了广泛的研究以寻找饲料中抗生素的天然替代品,并且已经发现植物化合物(或植物性化合物)具有巨大的潜力(Yang等,2015 )。其中,植物 单宁酸 受到了广泛关注,并且可能是研究最多的化合物,尤其是用于农场的化合物。 反刍动物。单宁的生物活性和动物对膳食单宁的反应已被广泛综述,主要侧重于动物的营养和生产(Mueller-Harvey等,2006; Waghorn,2008; Wang和McAllister,2011; Redondo等,2014)。在简要概述植物单宁的一些重要化学特性和生物学特性之后,本综述主要关注单宁在农场动物中的抗微生物,抗寄生虫和免疫调节特性的应用的最新进展。
2.化学结构和单宁的发生
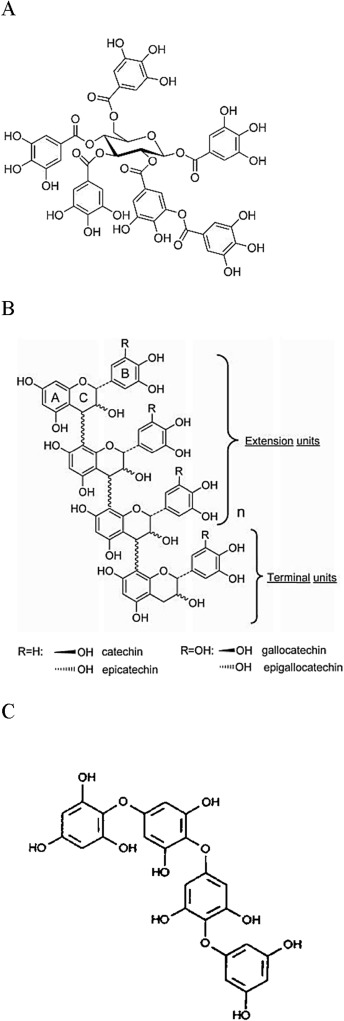
单宁 是天然存在的具有不同结构的酚类化合物的异质基团,它们具有结合和沉淀蛋白质的能力。单宁主要分为3大类: 可水解单宁 (HT), 缩合单宁 (CT)(也称为 原花青素)和间 苯单宁 (PT)。前两组位于陆生植物中,而PT仅存在于海生褐藻中(图1)。可水解单宁酸由 多元醇 核(通常为D-葡萄糖)组成,该多元醇核被酚酸(主要是没食子酸或六羟基二苯甲酸)酯化。HT的分子量为500至3,000 Da(Haslam,1989)。它们很容易被酸,碱或 酯酶水解,因此很容易降解并吸收到 消化道中, 并可能在食草动物中引起潜在的毒性作用(Dollahite等,1962; Mcleod,1974 )。缩合单宁是 由黄烷-3-醇单元(包括儿茶素, 表儿茶素,没食子儿茶素和 表没食子儿茶素)组成的低聚或聚合 类黄酮。 。与HT相比,CT具有更复杂的结构和1,000至20,000 Da的更高分子量。与HT不同,只有强的氧化和酸性水解作用才能使CT结构解聚,而CT结构也不易发生厌氧酶降解( McSweeney等,2001)。PT的结构不如陆地单宁(HT和CT)复杂,是间苯三酚(1,3,5-三羟基苯)的聚合反应形成的( Ragan和Glombitza,1986)。PT的分子量范围为126 Da至650 kDa( Targett and Arnold,1998),根据其化学结构,可分为6类(邻苯三酚,异氟烷,Echole,Fucole,Fuhalols,Fucophlorethols)。它们主要是通过乙酸-丙二酸途径合成的(Herbert,1989),尽管已经提出了另一种途径(Chen等,1997)。
{kind=link}
{kind=link}
图1。 (A)可水解单宁,(B) 缩合单宁 和(C) 邻苯二酚的模型结构 。
单宁在植物界广泛分布,尤其是在营养重要的草料,灌木,谷物和草药中含量丰富 (Salunkhe等,1982; Wang等,1999 )。它们还存在于许多水果种类中,例如香蕉, 黑莓,苹果和葡萄以及茶中(Nonaka等,1984; Bravo等,1992; Mertz等,2007; Mercurio和Smith,2008; Kheng ,2010年)。浓缩单宁是饲草豆科植物,树木和灌木中最常见的单宁类型, 而HT通常出现在树木的叶子中并在热带地区浏览灌木(Min等,2003)。通常,单宁在植物的脆弱部位,例如新叶和花朵中含量更高(Terrill等,1992; Van Soest,1982; Frutos等,2004 )。的PT都集中在位于所述physodes 细胞质 细胞的外皮质层内 菌体 (拉根和Glombitza,1986, Lüder和Clayton,2004, Shibata等人,2004)。单宁的化学结构和浓度在植物物种,生长阶段和生长条件(例如温度,光照强度,养分胁迫和接触草食动物)之间差异很大(Frutos等,2004)。, Amsler and Fairhead,2006; Berard等,2011; Li等,2014; Huang等,2017)。
3.单宁的生物活性
单宁 是植物的 次生代谢产物 ,可作为植物化学防御系统的一部分,抵抗病原体的入侵和昆虫的攻击。单宁酸已显示出许多生物活性,其中一些对现代食用动物的生产最重要。
3.1。抗菌性能
单宁的抗微生物活性早已被认识,并且单宁对细菌,真菌和酵母的毒性已得到综述(Scalbert,1991 )。迄今为止提出的解释单宁酸抗菌活性的机制包括抑制细胞外微生物酶,剥夺微生物生长所需的底 物,通过抑制氧化磷酸化对微生物代谢的直接作用 ,剥夺金属离子或与 细菌细胞膜形成复合物 。引起细胞壁形态变化并增加 膜通透性 (Scalbert,1991 ; Liu等,2013)。有证据表明,微生物细胞膜是单宁酸通过细胞聚集和破坏细胞膜及功能而抑制作用的主要部位(Mcallister等,2005; Liu等,2013)(图2 )。尽管蛋白质沉淀是所有单宁的普遍特性,但单宁的抗微生物活性是特定于微生物物种的,并且与单宁的化学组成和结构密切相关。通常,单宁酸对革兰氏阳性细菌的抗微生物活性 据报道要大于对 革兰氏阴性细菌的抗微生物活性 ( Ikigai等,1993; Smith and Mackie,2004)。),因为革兰氏阴性细菌具有由脂双层 结构组成的外膜,该双层结构由 脂多糖 和蛋白质的外层组成,而内层则由 磷脂组成 。然而,单宁特别是从几种植物中分离得到的CT已显示出对革兰氏阴性细菌具有很强的活性。值得注意的是,诸如 O157:H7大肠杆菌, 沙门氏菌, 志贺氏菌, 葡萄球菌, 假单胞菌 和 幽门螺杆菌等致病菌 均对单宁敏感(Funatogawa等,2004; Doss等,2009; Banso和Adeyemo,2010; Liu等,2013)。 Wang等。(2013年) 比较了12种单宁,发现仅从紫色草原三叶草(Dalea purpurea Vent)分离的CT 和从褐藻(Ascophyllum nodosum)分离的PT 具有强大的抗大肠杆菌 和抗大肠杆菌 O157:H7活性。 邻苯二酚的 抗菌活性也比CT和HT高(Wang等,2009)。研究表明,单宁酸氧化后的羟基数量和过氧化氢的释放是造成单宁酸抗菌性能的两个重要因素(Akagawa等,2003;Smith等,2003; Science等,2003。 Mueller-Harvey,2006 )。有人提出, 具有三羟基B环的黄酮醇(gallocatechin) 比 具有二羟基B环的儿茶素对链球菌, 梭菌, 变形杆菌 和 葡萄球菌具有更大的抑制作用 (Sakanaka et al。,1989 )。同样,表儿茶素没食 子酸酯和表没食子儿茶素没食子酸酯对肉毒梭菌的毒性 要大于未加盖的儿茶素梭菌素和 表没食子 儿茶素(Okuda等人,1985年)。)。由于单宁的来源广泛,从而导致其抗菌活性的巨大差异,因此筛选和鉴定对目标微生物有效且特异的单宁将一直是一项研究工作。
{kind=link}
{kind=link}
图2。 与(A)0(对照)或(B)200μg/ mL 紫色大草原三叶草的浓缩单宁酸(Dalea purpurea Vent。)孵育10 h的大肠杆菌(菌株25922)细胞的 聚集 ,以及E的透射电子显微照片 。大肠杆菌 O157:H7菌株3081与(C)0或50μg/ mL的(D)邻苯二酚,(E) 可水解单宁 或(F)缩合单宁孵育24小时 。改编自 Wang等,2009, Wang等,2013 和 Liu等。(2013)。
3.2。抗寄生虫性
单宁的抗寄生虫特性已 在体外 和 体内 研究中得到证实。 缩合单宁 从提取的 豆科 tanniferous 牧草 如 红豆草 (红豆 刺),大三叶(莲花长柄), 百脉根 (百脉根)和苏拉(岩黄芪 冠的)降低的比例 毛圆 蛇形毛圆线虫 孵化的卵和抑制卵发育 肺蠕虫 和 胃肠线虫 (混合物种 奥斯特线虫, 食道口线虫, 古柏, 毛圆,和 粪以剂量依赖的方式() Molan等人,2000年, Molan等人,2000b, Molan等人,2002 )。四种热带tanniniferous植物提取物已显示 驱虫 上效果 ħ aemonchus线虫 和 T. 蛇形毛圆线虫,主要干扰幼虫exsheathment的处理(阿隆索-Diaz等人。,2008年, 阿隆索-Diaz等人,2008年b)。栎树(Athanasiadou等,2001 ), 菊苣 (Molan等,2003)和绿茶(Molan等,2004)中的单宁提取物以剂量依赖的方式显着抑制幼虫的迁移。这些结果表明,单宁酸在寄生虫生命周期的不同阶段均具有抗寄生虫作用。 在许多 涉及绵羊和牛的体内研究中也已经证实了在体外研究中 观察到的各种单宁的抗寄生虫作用 (Paolini等人,2003; Heckendorn等人,2007; Chaweewan等人,2015; Desrues等人,2016年)。
植物单宁的驱虫机制已通过“直接”单宁对寄生虫细胞1降低主机建立感染第三期幼虫的从而降低了主机的入侵; 2)减少排泄作用)建议 线虫 的成虫鸡蛋蠕虫和3)减少卵向第三阶段幼虫的发育(Athanasiadou等,2001; Athanasiadou等,2005; Brunet等,2008; Hoste等,2012),并通过“间接”作用改善宿主对线虫的抗性(Coop和Kyriazakis,2001 ; Min等,2003 ; Tzamaloukas等。,2006 ; Pathak等,2016)。但是,与它们的抗菌活性相似,单宁的驱虫效果因单宁的化学组成和结构,寄生虫物种或生长阶段和/或宿主的物种而异(Hoste等,2006; Hoste等, 2012)。
3.3。抗氧化性能
长期以来,天然存在的酚类化合物一直被认为是有效的抗氧化剂(Rice-Evans等,1995; Rice-Evans等,1996 )。单宁的抗氧化特性已广泛用于食品工业和医学领域,以预防 与氧化应激相关的疾病,如心血管疾病,癌症或骨质疏松症(Hollman和Katan,1999; Scalbert等,2005 )。研究表明,相对高分子量的CT和HT表现出 比简单酚类更高的 抗氧化活性(Hagerman等,1998)。)。羟基的数量和单宁的聚合度被认为与它们清除自由基的能力有关(Ariga和Hamano,1990)。具有最多羟基的单宁酸最容易被氧化(Hodnick等,1988),因此具有最大的抗氧化活性。 Ricci等。(2016年) 表明,单宁酸作为天然抗氧化剂的有效性归因于其还原和氧化还原活性的复杂组合,这也有助于其清除自由基的能力。
单宁酸作为生物抗氧化剂的潜力已在许多 体外 研究中表明(Ho等,1999; Lin等,2001; Beninger和Hosfield,2003; Barreira等,2008)。在 体内 的单宁的抗氧化活性也显示出在不同的 动物组织。包含CT的草料的加入通过增加血清抗氧化活性来改善牛和羊的抗氧化状态(Dutta等,2012 ; Dey和De,2014 ; Huang等,2015 ; Peng等,2016)。羔羊日粮中的Quebracho单宁改善了肌肉(Luciano等,2011),肝脏和血浆的抗氧化状态(López-Andrés等,2013 ),并通过延迟 冷藏过程中的肌红蛋白氧化来增强了肉的颜色稳定性 (Luciano等。 ,2009年)。鉴于HT在吸收前已在胃肠道中降解,而CT无法从消化道降解和吸收 ,因此不容易解释单宁如何作为完整实体在动物体内发挥抗氧化活性。 López-Andrés等。(2013年) 发现quebracho单宁酸不会在胃肠道中降解或吸收,但会增加绵羊肝脏和血浆的抗氧化能力,这表明CT可能间接影响动物组织的抗氧化状态。单宁蛋白复合物已显示出减少但不能消除单宁的抗氧化活性(Riedl和Hagerman,2001; Arts等,2002)。据推测,膳食单宁可以在消化过程中保留其他营养抗氧化剂,或者可以保护消化道中的蛋白质,碳水化合物和脂质免受消化过程中的氧化损害(Marshall和Roberts,1990年))。但是,单宁在动物组织中的抗氧化机制尚不清楚。需要在这一领域进行进一步的研究,特别是因为提高抗氧化剂的水平被认为是单宁喂养给动物福祉和性能带来的最大好处之一。
3.4。抗炎特性
单宁具有不同 的抗炎活性 (Mota等,1985; Terra等,2007; Sugiura等,2013; Park等,2014),与它们的抗氧化活性呈正相关(Gonçalves等, 2005年, Souza等人,2007年, Park等人,2014年)。 体外 研究表明,葡萄籽中的单宁酸 通过调节细胞因子的表达降低了肥胖等低度炎症性疾病(Terra等,2007; Chacón等,2009)。)。从黑树莓种子中提取的CT的抗炎活性通过抑制脂多糖诱导的RAW 264.7细胞产生一氧化氮(NO)的能力得到证明,NO是一种诱导炎症的促炎介质(Park等,2014 )。 可水解单宁 从 水柏枝苞 显示对一个显著抗炎作用 巴豆 小鼠和在DBA / 1小鼠胶原诱导的关节炎(油诱导的耳水肿Liu等人,2015A, Liu等人,2015B)。作者推测,抗炎作用的机制与清除自由基的有效能力有关,而不是与HT对NO和促炎性细胞因子产生的抑制作用有关。从鼠尾藻多酚 A.结节性红斑 和 Ecklonia 静脉 也显示出基于它们的能力,以抑制细胞因子释放有效的抗炎效果(Dutot等人,2012 ),NO和 前列腺素E2 制作(Wijesinghe等人,2013)。需要指出的是,该领域的大多数研究都是使用 体外 模型进行的。单宁在消化后在动物体内的抗炎作用的功效需要进一步评估。 体内 模型。
3.5。防病毒属性
单宁已显示出对某些病毒的显着活性,例如 人类免疫缺陷病毒 (HIV),牛腺相关病毒和 诺如病毒 (Uchiumi等,2003; Di Pasquale等,2012; Zhang等,2012)。)。 杨等。(2013) 发现HT(chebulagic acid)在体外具有相当大的抗肠病毒71活性 , 并通过抑制小鼠模型中的病毒复制有效降低了死亡率并缓解了临床症状。已经证明,单宁通过靶向病毒逆转录酶来抑制HIV-1复制,从而对HIV-1产生抑制作用( Tan等,1991)。),蛋白酶(Xu等,2000)和整合酶(Au等,2001 ),或通过干扰gp41性螺旋束的形成来抑制HIV-1进入靶细胞(Liu等,2004)。 。据报道,绿茶中的表没食子儿茶素抑制 丙型肝炎病毒 (HCV)的进入(Ciesek等,2011; Calland等,2012)。 Liu等人,2015a, Liu等人,2015b 揭示了 单宁酸 (HT)抑制HCV进入和细胞间传播,但不干扰细胞内HCV复制。三种HT(punicalagin,punicalin和geraniin)抑制 乙型肝炎病毒 通过防止cccDNA的形成和促进cccDNA衰变的双重机制产生cccDNA(Liu等人,2016)。 上田等。(2013) 发现柿子中的单宁酸 (Diospyros kaki)显着降低了12种被测试病毒的病毒感染性,而绿茶,相思树 和花生仁中的单宁酸 仅对其中一些有效,蛋白质聚集似乎是抗病毒的基本机制。 -柿单宁的病毒作用。 已证明从大肠杆菌中分离出的邻苯二酚 具有抗流感病毒神经氨酸酶的强活性(Ryu等人,2011年),猪流行性腹泻病毒(PEDV)通过抑制病毒进入和/或病毒复制(Kwon等人,2013)和HIV-1(Karadeniz等人,2014)。类似的抗病毒活性也被证明为PT分离自 赤子爱胜 bicyclis 针对鼠诺罗病毒(严等人,2015)和人乳头状瘤病毒(Kim和郭,2015年)。
以上所有信息表明,单宁具有不同的抗病毒活性,具体取决于化学成分和结构。尽管单宁酸的抗病毒作用方式尚不完全清楚,但到目前为止获得的信息表明,抑制病毒吸附到细胞中,抑制病毒渗透到 细胞核中 以及抑制病毒逆转录酶可能是一些潜在的机制。 (Buzzini et al。,2008)。 需要进行体内研究以探索单宁酸作为天然抗病毒剂在动物和家禽行业中使用的潜力。
4.在反刍动物中使用单宁酸
单宁酸 特别是CT广泛分布在具有重要营养价值的 饲草,树木,灌木和 豆类中,反刍动物通常会食用它们 。因此,CT对 反刍动物营养,健康和生产的影响已被广泛研究和综述(Frutos等,2004; Mueller-Harvey,2006; Waghorn,2008; Patra和Saxena,2011; Wang等,2015)。 。 单宁浓缩 可以对反刍动物产生有益或有害的影响,这取决于反刍动物的食用量,其类型和化学结构以及其余日粮的组成,尤其是日粮中的CP浓度(Mueller-Harvey,2006年)。人们普遍认为,在中低浓度(<50 g / kg DM)浓度的温草料中,CT可以提高反刍动物的蛋白质利用率,而又不会对饲料的摄入和营养物质的消化产生不利影响(Barry和Mcnabb,1999; Waghorn,2008 ),取决于CT来源和用于确定浓度的分析方法/标准。蛋白质沉淀能力,抗微生物,抗寄生虫和 抗氧化活性 单宁在反刍动物中的用途是最相关的特性。通过总结大量研究, Waghorn(2008) 的结论是,当饲料被馈送作为唯一饮食中,CT在 L.百脉 (约30g CT /千克DM)已经用于反刍生产有利,但在CT 红豆草, ħ edysarum coronarium 和 L. pedunculatus (浓度通常大于50 g / kg DM)除了减轻寄生虫的影响外,似乎没有对生产力产生任何好处。与温带耕作相反,在温暖和炎热的气候中,典型的浏览CT几乎总是有害于反刍动物,除了减少内部寄生虫的数量(Waghorn,2008 )。中低浓度的浓缩单宁酸有利于反刍动物的生产效率,因为CT可以减少瘤胃中蛋白质的降解 ,增加 日粮蛋白质 到达小肠吸收的量( Wang等,1994; Wang等,1996 )。然而,在高浓度下,CT由于其涩性而会阻碍饲料的摄入,并通过“过度”保护蛋白质来减少蛋白质和其他营养物质的消化,降低瘤胃 微生物活性 并抑制内源性消化 酶的活性 从而对动物的表现产生负面影响。饮食浓度CT对动物的生产性能产生负面影响,这再次取决于CT的来源(即化学成分和结构)。因此,大多数研究集中于筛选和评估不同单宁来源,并确定反刍动物日粮中它们的最佳浓度。这些包括筛选和鉴定潜在的含单宁-饲料(例如, L.百脉, L.长柄,红豆草),其可以是合并到动物饮食( Barry和麦克纳布,1999, 贝拉尔等人,2011, 阿查等。 ,2013),以定义各种外部单宁(例如quebracho单宁,单宁酸)对动物生长性能的最佳补充率(Dschaak等人,2011 ; Anantasook等人,2015 ; Rivera-Méndez等人,2017)),并在集约化饲喂操作中发展技术(例如碱处理,单宁结合剂,饮食混合),以在热带和其他饲料来源受限的地区使用富含单宁的树叶和灌木(Murdiati等, 1990, Smith等,2005, Wina等,2005, Brown和Ng’Ambi,2017)。
单宁酸在反刍动物生产中最成功的应用可能是减少泡沫膨胀。膨胀是反刍动物中常见的消化系统疾病。该病的特征是瘤胃和网状结构 中积聚了气体 ,可能损害消化功能和呼吸功能(Wang等,2012 )。许多因素可以向膨胀,但植物细胞并在其进入瘤胃增加的粘度从植物细胞中蛋白质的释放,迅速裂解 瘤胃液 是牧场膨胀的主要因素。单宁酸通过在咀嚼和反刍过程中沉淀蛋白质来降低蛋白质在瘤胃中的溶解度,从而减少膨胀的发生。因此,含单宁的饲料被认为是“无膨胀的”。 Li等。(1996年) 估计,仅需1.0 mg CT / g DM即可预防牧场膨胀。 已证明将含CT的牧草如皂角苷混入 苜蓿中是控制苜蓿牧场膨胀的有效方法(图3; Wang等人,2006; Sottie等人,2014)。
{kind=link}
{kind=link}
图3。 1999年和2000年 确定的for牛(n = 12)放牧 牧草重新生长所测定的草量中mass 肿率和皂甙的比例回归 (改编自 Wang等人(2006))。
单宁酸在反刍动物中特别是在放牧反刍动物中的另一个主要应用是控制消化寄生虫。 Hoste等。(2006) 总结 了体内 研究,表明皂素,苏拉,pedunculatus, Sericea lespedeza, Acacia nilotica 和 菊苣中的单宁酸 对绵羊,山羊和鹿的消化道 有明显的 驱虫 作用 。外部单宁,例如来自含羞草 (HT),栗子(HT)和quebracho(CT)的单宁, 已被用来控制反刍动物中的各种肠道寄生虫( Butter等,2001; Min and Hart,2003, Min等人,2005, Min等人,2015)。膳食中单宁酸浓度低于20 g / kg DM似乎不能有效控制反刍动物肠道寄生虫。一个有趣的现象是绵羊可以检测到内部寄生虫或相关症状的存在,并增加了它们对富含单宁的饲料的偏好(Lisonbee等,2009; Juhnke等,2012)。
近来,已经研究将单宁或含单宁的饲料掺入饮食中作为减少反刍动物消化道中食源性病原体的收获前方法(表1)。紫色草原三叶草已被证明可以缓解 大肠杆菌 O157:H7,这是一种致命的食源性致病细菌,位于反刍动物消化道中(Jin等,2015; Huang等,2015)。 黄等。(2015) 发现,与挑战羔羊 大肠杆菌 O157:H7喂食含有36克紫色草原的三叶草CT / kg的DM棚显著较少的饮食 大肠杆菌 O157:H7比饲喂没有CT的小羊。同样的CT也被证明可以降低 放牧天然草场的牛的大肠杆菌粪便脱落含量为16至20 g CT / kg DM(Jin等,2015)。另外,以15克/千克DM的浓度添加鞣酸栗降低粪便脱落 大肠杆菌 用于 馈送牛 干草日粮(Min等人,2007)。然而,其他单宁酸,如 较低浓度(<13.5 g CT / kg DM)中的皂甙和沙雷氏菌( S. lespedeza),对粪便大肠杆菌的 脱落没有影响 (Lee等,2009a; Lee等,2009b; Berard)等人,2009)。结节曲霉的补充 日粮中浓度为10至20 g / kg DM的粗粉可显着降低 牛的大肠杆菌 O157:H7粪便脱落(表1)。这些研究中未报告单宁浓度,但 结节曲霉 是褐藻,含有高水平的PT(Wang和McAllister,2011)。这些研究表明,饲喂植物单宁可以作为一种有效的方法,以有效减少 反刍动物消化道中大肠杆菌 O157:H7 的存在, 从而降低car体污染的风险,从而提高食品安全性。
表1 。饮食的影响 单宁 对 大肠埃希氏菌和大肠杆菌中 O157:H7的粪便脱落 反刍动物。
|
资料来源 |
单宁类型 |
动物 |
饮食 |
申请率 |
观察效果 |
参考文献 |
|
泡叶 藻 提取物 |
芦丁宁(PT) |
育肥牛 |
谷物饮食 |
屠宰前14天,每公斤体重为0、10或20克 |
减少粪便和生皮 大肠杆菌 和 大肠杆菌 O157:H7 |
|
|
A. 结节 餐 |
PT |
牛 |
谷物饮食 |
20克/千克DM |
减少粪便和生皮 大肠杆菌 O157:H7和 沙门氏菌 |
|
|
A. 结节 餐 |
PT |
育肥牛 |
玉米饮食 |
日粮20 g / kg,屠宰前14天 |
降低 E. 大肠杆菌 O157:上隐藏拭子和粪便样品中H7患病 |
|
|
A. 结节 餐 |
PT |
大肠杆菌 O157:H7挑战牛 |
大麦浓缩饮食 |
10或20 g / kg的饮食,最多14天 |
减少粪便中大肠杆菌 O157:H7的脱落 |
|
|
紫色草原三叶草 |
浓缩单宁(CT) |
大肠杆菌 O157:H7挑战羔羊 |
新鲜草料 |
36克/千克DM ECT |
减少 大肠杆菌 O157:H7粪便脱落 |
|
|
芹ice |
电脑断层扫描 |
山羊 |
有 |
13.5克/千克DM ECT |
对粪便大肠杆菌无影响 |
|
|
板栗 |
可水解单宁(HT) |
牛 |
有 |
15克/天 |
减少粪便 大肠杆菌 |
|
|
皂素 |
电脑断层扫描 |
牛 |
干草/青贮 |
1.1至12 g / kg DM |
对粪便脱落没有影响 |
|
|
紫色草原三叶草 |
电脑断层扫描 |
牛 |
新鲜草料 |
16至20 g / kg DM ECT |
减少粪便 大肠杆菌的 脱落 |
ECT =可萃取的 浓缩单宁。